Введение
Засуха характерна для территорий с аридным климатом. Поскольку она оказывает негативное влияние на состояние экосистем, ежегодно из-за засухи отмечается увеличение неиспользуемых сельскохозяйственных угодий, площади деградированных территорий и наносится значительный ущерб экономике по всему миру (Hermans, McLeman, 2021; Pugacheva, 2023). Борьба с засухой является одной из важнейших задач (Feeley et al., 2020; Yan et al., 2022; Eldridge et al., 2024). Для этого применяются агролесомелиоративные мероприятия, в частности, поиск и высадка видов древесных растений, устойчивых к засухе и обладающих высокой биологической продуктивностью (Kulik et al., 2023). Еще одним важным способом XXI века в условиях климатических изменений, вызванных природными и антропогенными факторами и способствующих увеличению CO2 в атмосфере, является отбор древесных растений с высоким ассимиляционным потенциалом, а также с высокой устойчивостью к засухе, способностью поддерживать высокую биологическую продуктивность в условиях аридного или субаридного климата (Javaid et al., 2022; Kurganova et al., 2022).
Одним из таких древесных растений является дуб черешчатый, Quercus robur L., который активно используется в агролесомелиорации и защитном лесоразведении юга России (Mikhin, Mikhina, 2019; Semerikova et al., 2023; Zamolodchikov et al., 2023). Однако молекулярно-генетические механизмы формирования высокой устойчивости к засухе и фотосинтетической продуктивности у Q. robur, произрастающего в условиях аридного климата слабо изучены. Это может быть связано с тем, что первый автоматически аннотированный референсный геном Q. robur появился только в 2022 г.
Одним из наиболее изученных ферментов, продукт которого участвует в формировании устойчивости к засухе у растений, является абсцизная альдегидоксидаза (AAO), катализирующая окисление абсцизового альдегида до абсцизовой кислоты (ABA). ABA, в свою очередь, активирует различные защитные механизмы растений, включая реакцию на засуху. Например, регулируя открытие и закрытие устьиц, что помогает растениям минимизировать потерю воды в условиях дефицита влаги (Trivedi et al., 2016; Chen et al., 2020). Другим не менее важным ферментом, обеспечивающим ассимиляцию CO2 и запускающим цикл Кальвина, является рибулозо-1,5-бисфосфаткарбоксилаза/оксигеназа (RuBisCO), состоящая из больших и малых субъединиц (Zhao et al., 2024). Эффективность работы RuBisCO напрямую влияет на фотосинтетическую продуктивность и, следовательно, на выживаемость растения в стрессовых условиях. Активность RuBisCO регулируется различными факторами, включая концентрацию CO2 (Carmo-Silva, Sharwood, 2023). Большая субъединица RuBisCO, кодирующаяся геном rbcL содержит активные центры, где происходит фиксация CO2 (Andersson, Backlund, 2008). Ранее (Galka et al., 2015) были получены интересные результаты, где предполагается, что ABA может связываться с RuBisCO и ингибировать его работу, но это требует дальнейшего изучения и утверждать это преждевременно. В исследовании (Ikegami et al., 2009) у Arabidopsis thaliana L. было показано, что в условиях засухи увеличивается уровень ABA в тканях растения. Аналогичные результаты были показаны у риса (Shi et al., 2021). Исходя из вышесказанного, можно предположить, что продукт гена ААО и функционирование RuBisCO связаны между собой, но у разных растительных объектов в стрессовых условиях это взаимодействие и его результат могут различаться.
Регуляция экспрессии генов осуществляется посредством связывания транскрипционных факторов с регуляторной последовательностью ДНК, тем самым стимулируя транскрипционную активность генов. Среди таких транскрипционных факторов выделяется семейство DREB, которое является важным элементом в регуляции реакции растений на стрессовые условия (Fujita et al., 2011), включая засуху (Rini, 2019; Hrmova, Hussain, 2021; Manna et al., 2021; Singh, Chandra, 2021). Существуют исследования, в которых изучалась связь между генами rbcL, DREB2A и AAO3. Например, применение экзогенной ABA улучшало фотосинтетическую работу редкого вида деревьев Emmenopterys henryi Oliv. за счет повышения активности RuBisCO в условиях стресса (Cai et al., 2023). Было показано, что DREB2 в основном активируется ABA-независимыми регуляторными путями в ответ на осмотический стресс и изменения температуры (Lata, Prasad, 2011). В то же время у A. thaliana факторы транскрипции других семейств генов, связанных с ABA, могут напрямую связываться с промотором DREB2A, тем самым повышая его транскрипционную активность и обеспечивая устойчивость растений к засухе (Kim et al., 2011). Аналогичные данные были получены и на пшенице (Egawa et al., 2006).
Содержание различных пигментов и вегетационные индексы являются хорошими индикаторами для оценки состояния растений при действии стрессоров. Например, в начале засухи наблюдается тенденция к увеличению содержания хлорофилла, а в последствии происходит его снижение (Nikolaeva et al., 2010). Согласно другим исследованиям, при засухе содержание флавонолов и полифенолов может повышаться (Diethelm et al., 2022). В ответ на стресс растения часто начинают синтезировать больше антоцианов. Это связано с их способностью защищать клетки от окислительного стресса, вызванного активными формами кислорода, которые накапливаются при дефиците воды. Индекс азотного баланса (NBI) отражает соотношение содержания хлорофилла и флавонолов, характеризуя азотное обеспечение растений (Chen et al., 2022; Jan et al., 2024). При засухе NBI обычно снижается из-за уменьшения содержания хлорофилла и увеличения синтеза флавонолов, что указывает на ухудшение азотного питания и активацию защитных механизмов.
Резюмируя вышесказанное, комплексное исследование паттернов экспрессии данных генов у Q. robur имеет важное значение для расширения представлений о молекулярно-генетических механизмах устойчивости древесных видов к засухе. В связи с этим целью исследования стало изучение реакций Q. robur на засуху путем оценки морфологических, физиологических параметров и транскрипционной активности генов.
Материалы и методы
Желуди были собраны с Q. robur, произрастающих на семенной плантации ФНЦ агроэкологии РАН (N48.632932, E44.419876). Сеянцы Q. robur выращивались в кассетах с универсальным грунтом и песком в соотношении 3 к 1. После этого были отобраны 30 особей (высота растений 10-15 см, c 4 и более настоящими листьями) и пересажены в отдельные горшки с тем же составом грунта. Сеянцы Q. robur были распределены на две экспериментальные группы по 15 растений в каждой. Влажность почвы измерялась каждый день утром с помощью влагомера MC-7828 SOIL (Landtek, Тайвань). В течение эксперимента в контрольной группе поддерживалась влажность почвы не менее 75-80% НВ, а в группе засуха – не более 10-15% НВ. Через одну и две недели после начала эксперимента проводилось измерение морфологических и физиологических параметров, а также отбор листовых пластинок для выделения РНК в контрольной (первая неделя: К-1, вторая неделя: К-2) и экспериментальной (первая неделя: З-1, вторая неделя: З-2) группах. Собранный материал хранился в криопробирках со стабилизатором IntactRNA (Евроген, Россия) при температуре -80˚С.
В качестве морфологических показателей оценивалась степень увядания листьев и высота растений. Для оценки степени увядания листьев Q. robur использовалась 6-балльная шкала, описанная в работе (Engelbrecht, Kursar, 2003). Высота надземной части растений измерялась с помощью линейки (см). Для оценки физиологических параметров проводилось неинвазивное измерение содержания хлорофиллов (мкг/см2 сырой массы, от 0 до 150), флавонолов и антоцианов (индексы с диапазоном значений от 0 до 3), индекса азотного баланса (NBI, от 0 до 999) в листовых пластинках Q. robur с помощью анализатора растений Dualex Scientific+TM («Force-A», Франция)(Shi et al., 2024).
Для выделения тотальной РНК из листовых пластинок Q. robur использовался набор R-Plants (Биолабмикс, Россия). Навески листьев Q. robur по 50 мг гомогенизировали с помощью Precellys® 24 (Bertin Technologies, Франция). Выделение и центрифугирование РНК осуществлялось при +4°С. Далее образцы тотальной РНК обрабатывались ДНКазой I (Magen, Китай) с последующим переосаждением в 3M ацетате натрия и этаноле. Концентрация РНК измерялась с помощью набора Equalbit RNA BR Assay Kit (Vazyme, Китай) на флуориметре Qubit® 4 (Thermo Fisher Scientific, США). Качество выделенной РНК оценивалось спектрофотометрически по соотношениям длин волн А260/A280 и A260/A230 с помощью SPECTROstar Nano (BMG Labtech, Германия), а также гель-электрофорезом в 1,2 % агарозном геле с использованием буфера для нанесения РНК на гель ФриК (Биолабмикс, Россия), содержащего формамид. Аликвоты выделенной РНК хранились при -80°С и подвергались только одному циклу замораживание-оттаивание перед ОТ-ПЦР-РВ.
Для анализа относительной экспрессии генов Q. robur была применена ОТ-ПЦР-РВ. Праймеры к целевым генам DREB2A-like, rbcL и AAO (к общему участку у обеих изоформ), подобранные с помощью онлайн-сервиса Primer-BLAST (NCBI), представлены в таблице 1. Референсный ген actin 7 был подобран ранее в наших исследованиях (Zybinskaya, Tretyakova, 2024).
Таблица 1 – Праймеры, использовавшиеся для ОТ-ПЦР-РВ
|
Ген |
GeneBank ID |
Прямой праймер (5’→ 3’) |
Обратный праймер (5’→ 3’) |
|
DREB2A |
XM_050409186.1 |
GAAAGAGGCTTTGGCTTGGC |
GTGGCAGTAGACTCATCAACCG |
|
rbcL |
AB125025.1 |
CTTGAATGCTACTGCAGGTA |
ATCCCGGCAATAATGAGC |
|
AAO |
XM_050428960.1 |
CTCAGGCCACTCAGGGAAAG |
GCGGCACCATAGGTTCGATA |
|
actin 7 |
XM_050436545.1 |
CGGAGGTTGGCTTGCTCTT |
ATCCAGCCTTCACCATTCCAG |
ОТ-ПЦР-РВ проводилась в амплификаторе Gentier 96E (Tianlong, Китай) с использованием набора OneTube RT-PCR SYBR (Евроген, Россия) с 50Х LowROX (Евроген, Россия) в качестве референсного красителя. Концентрация каждого образца РНК была приведена к 20 нг/мкл путем разбавления. Общий объем реакционной смеси составлял 25 мкл, содержащий 5 мкл 5X OneTube PCRmix SYBR, по 1 мкл каждого праймера (10 мкМ), 0,5 мкл 50Х LowROX, 2 мкл РНК, 0,5 мкл ревертазы и 15 мкл воды, свободной от нуклеаз. Для каждого образца реакция проводилась в трех технических проворностях. Протокол ОТ-ПЦР-РВ был следующим: обратная транскрипция при 55°С в течение 15 мин, активация полимеразы и инактивация ревертазы при 95°С в течение 1 мин, далее денатурация при 95°С в течение 15 с, отжиг праймеров при 62°С (actin 7 и rbcL) или 64°C (AAO и DREB2A) в течение 20 с, элонгация при 72°С в течение 20 с, всего 50 циклов и построение кривой плавления при 58-95°C с шагом 0,5°С. Значения пороговых циклов амплификации (Ct) были получены автоматически. Относительная экспрессия целевых генов была рассчитана по методу 2–∆∆Ct (Livak, Schmittgen, 2001).
Количественные данные обрабатывались с помощью программы Statistica 12.0 (StatSoft Inc., 2020) с расчетом показателей, принятых для характеристики непараметрических выборок, поскольку данные относительной экспрессии имели ненормальное распределение: медиана, 1-й квартиль, 3-й квартиль (Me [Q1 ÷ Q3]). Для оценки значимости различий между контрольными и экспериментальными группами был использован тест Манна-Уитни для двух независимых выборок (p <0,05), а между первой и второй неделей – критерий Вилкоксона (p <0,05). При проверке статистических гипотез при анализе экспрессии трех генов в контрольной и экспериментальной группах был применен ANOVA Фридман тест (p <0,05), для post hoc анализа двух парных выборок – критерий Вилкоксона с поправкой Бонферрони (p <0,017) (Grjibovski et al., 2016). Для оценки корреляционной зависимости значений использовали метод ранговой корреляции Спирмена. Корреляция оценивалась как слабая (r <0,5), умеренная (r в интервале от 0,5 до 0,8) и сильная (r >0,8) при значимости p <0,05.
Результаты исследования
В течение эксперимента рост растений Q. robur из контрольной группы (табл. 2) увеличился в 1,17 раза (p = 0,001). Без полива первые признаки увядания (небольшая потеря тургора, изменение угла наклон листьев) наблюдались уже через одну неделю (З-1), а через две недели (З-2) отмечали изменение структуры поверхности листьев с начинающимся некрозом у отдельных растений. Оценка степени увядания листьев Q. robur выявила статистически значимые различия между экспериментальной и контрольной группами. Через одну неделю степень увядания листьев у З-1 была выше на 1 балл по сравнению с К-1 (p = 0,020), а на второй неделе между З-2 и К-2 разница составила 2 балла (p = 0,001).
Таблица 2 – Морфологические и физиологические параметры Q. robur

* – статистически значимые различия с контрольной группой (критерий Манна-Уитни, при p <0,05) и # – с той же группой на другой неделе эксперимента (критерий Вилкоксона, при p <0,05); Me – медиана, Q1 и Q3 – первый и третий квартили соответственно.
Содержание хлорофилла (табл. 2) в листьях исследуемых растений в условиях полива снизилось у К-2 по сравнению с К-1 (p = 0,038). Кроме того, оно было выше в 1,15 раз у З-2 по сравнению с К-2 (p = 0,005), но на первой неделе эксперимента между З-1 и К-1 различий не было выявлено. Содержание флавонолов на второй неделе эксперимента у З-2 повысилось по сравнению с З-1 (p = 0,038). Кроме того, содержание флавонолов отличалось между З-1 и К-1 (p = 0,018) и З-2 и К-2 (p = 0,032). В целом, растения из групп с поливом и без полива продемонстрировали различную реакцию (К – снижение, З – рост). Содержание антоцианов в листьях растений З-2 повысилось по сравнению с З-1 (p = 0.009). В то же время, оно было меньше у З-1 по сравнению с К-1 и у З-2 по сравнению с К-2 (p = 0,026 и p = 0,023 соответственно). Таким образом, как при наличии, так и при отсутствии полива растения отреагировали повышением содержания антоцианов в листьях. Значения NBI у растений из группы З-1 были выше по сравнению с К-1 (p = 0,001). Кроме того, у З-2 они снизились по сравнению с З-1 (p = 0,039) до уровня группы К-1. В целом, у контрольной и экспериментальной групп наблюдалось снижение значений NBI в ходе исследования.
Таким образом, в контрольных и засушливых условиях исследуемые сеянцы Q. robur демонстрировали схожую реакцию: для них было характерно повышение высоты надземной части, степени увядания листьев и содержания антоцианов; снижение содержания хлорофиллов и значений NBI. Различия в реакциях были обнаружены только по содержанию флавонолов: в отличие от контроля, при засухе оно повышалось.
Анализ результатов ОТ-ПЦР-РВ у Q. robur показал, что растения с поливом и без полива имели противоположную реакцию на уровне экспрессии генов DREB2A, rbcL и AAO. Экспрессия DREB2A была максимальной у З-1 (рис. 1а), по сравнению с К-1 она была выше в 2,7 раза (p = 0,001). На второй неделе эксперимента у группы без полива (З-2) произошло снижение экспрессии DREB2A до уровня К-1 (p = 0,043), а у К-2 экспрессия, напротив, незначительно повысилась. Таким образом, отсутствие полива у З-1, З-2, а также с полив у К-1, К-2 оказало влияние на экспрессию DREB2A на разных сроках эксперимента: у З – на первой неделе, у К – на второй.
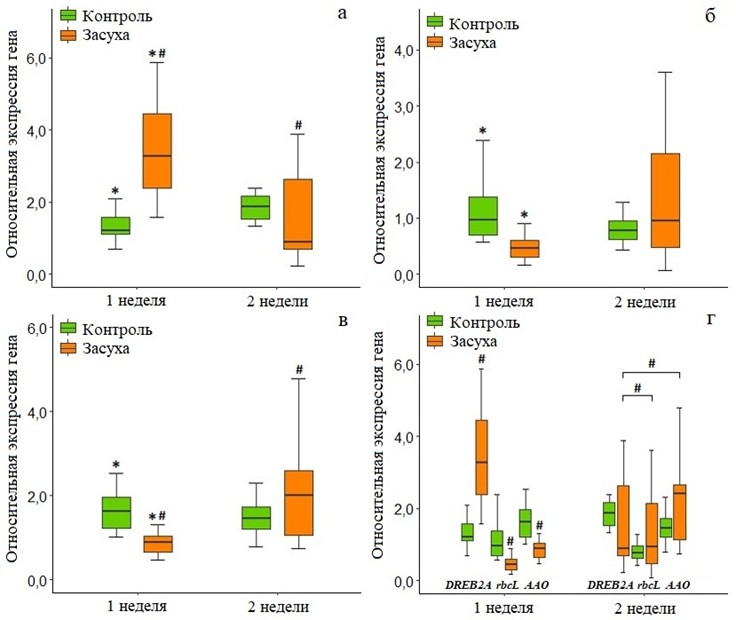
Рис. 1 – Относительная экспрессия генов DREB2A, rbcL и AAO у контрольной и экспериментальной группы Q. robur (a – DREB2A; б – rbcL; в – AAO; г – сравнение между генами DREB2A, rbcL и AAO). * – статистически значимые различия по критерию Манна-Уитни (p <0,05), # – по критерию Вилкоксона (p <0,05, для рисунка Г – p <0,017)
Противоположные DREB2A тенденции наблюдались при оценке экспрессии rbcL (рис. 1б): у контрольной группы она незначительно снижалась к концу второй недели (К-2), у экспериментальной (З-2) – повышалась. Кроме того, у З-1 экспрессия rbcL была ниже по сравнению с К-1 (p = 0,001), но к концу второй недели восстановилась до уровня контроля. В целом, растения из группы без полива отреагировали сначала снижением уровня экспрессии rbcL относительно контроля на первой неделе эксперимента (З-1), а к концу второй недели (З-2) – повышением. Реакция растений из группы с поливом была незначительной – небольшое снижение экспрессии rbcL к концу второй недели (К-2).
Схожая с rbcL реакция Q. robur наблюдалась при оценке уровня экспрессии AAO (рис. 1в): незначительное снижение к концу второй неделе у К-2 и рост в 2,7 раз (p = 0,002) у З-2 по сравнению с З-1. Помимо этого, у З-1 по сравнению с К-1 уровень экспрессии AAO был ниже (p = 0,001), но у З-2 превысил уровень контроля. Таким образом, отсутствие полива привело у группы З сначала к снижению, а затем росту уровня экспрессии гена AAO выше уровня контроля. Контрольная группа растений с повышенной температурой и поливом отреагировала небольшим снижением уровня экспрессии этого гена на второй неделе эксперимента.
Сравнение уровней экспрессии DREB2A, rbcL и AAO внутри каждой группы (рис. 1г) выявило наличие различий у З-1 и З-2. При попарном сравнении у З-1 были обнаружены различия в экспрессии трех изученных генов: DREB2A и rbcL (p = 0,001), DREB2A и AAO (p = 0,001), rbcL и AAO (p = 0,001). Наибольшим уровнем транскрипционной активности у З-1 обладал ген DREB2А, по сравнению с rbcL и AAO его экспрессия была выше в 7,2 и 3,7 раза соответственно. В группе З-2 попарное сравнение выявило различия в уровнях экспрессии между следующими генами: DREB2A и rbcL (p = 0,015), DREB2A и AAO (p = 0,015). В целом, у растений в условиях дефицита влаги на первой неделе эксперимента (З-1) преобладала экспрессия гена DREB2A, а на второй неделе (З-2) она снизилась до уровня контроля и преобладающей стала экспрессия AAO.
Результаты оценки транскрипционной активности генов, а также измерения роста и содержания пигментов в листовых пластинках Q. robur были проанализированы на наличие корреляций между исследуемыми показателями у контрольной и экспериментальной групп (рис. 2).
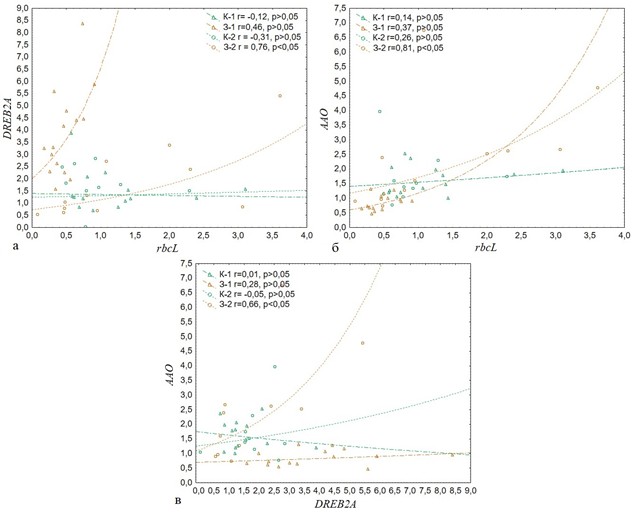
Рис. 2 – Результаты корреляционного анализа относительной экспрессии генов Q. robur (a – DREB2A и rbcL; б – AAO и rbcL; в – AAO и DREB2A в контрольной (К-1 и К-2) и экспериментальной (З-1 и З-2) группах
Группа З-2 имела умеренную положительную корреляцию между уровнями транскрипционной активности генов DREB2A и rbcL (рис. 2а), DREB2A и AAO (рис. 2в), а также сильную положительную корреляцию между кbcL и AAO (рис. 2б). Относительная экспрессия rbcL имела умеренную отрицательную корреляцию (-0,64) с содержанием антоцианов в группе З-1. Также, была выявлена умеренная отрицательная корреляция между уровнем экспрессии DREB2A и содержанием хлорофилла у растений З-2 (-0,59).
Обсуждение
Механизмы защиты от стресса действуют на различных уровнях: молекулярно-генетический, клеточный, тканевый и т д. (González, 2023; Qiao et al., 2024; Sabouri et al., 2024). В нашем исследовании мы наблюдали изменение экспрессии DREB2A, rbcL и AAO, а также содержания метаболитов в листовой пластике в условиях моделируемой засухи у Q. robur.
Одним из важных индикаторов жизненного состояния растений может быть содержание различных пигментов, например, хлорофилла, по которому можно судить об активности фотосинтеза и в целом об уровне стресса, испытываемого растением (Carter, Knapp, 2001). В нашем исследовании мы наблюдали снижение содержания хлорофилла в листьях Q. robur как у контрольной, так и экспериментальной групп. Полученные нами результаты согласуются с некоторыми ранее проведенными исследованиями влияния засухи на содержание хлорофиллов у других видов рода Quercus, например, Q. brantii (Jafarnia et al., 2018). В то же время в другом исследовании у Q. pubescens (Gallé et al., 2007) содержание хлорофиллов в листьях практически не изменялось во время засухи.
Еще одним важным классом пигментов являются антоцианы, высокое содержание которых может быть индикатором того, что растение находится в стрессовом состоянии, в том числе из-за засухи (Gould et al., 2002). В нашем эксперименте мы наблюдали схожие изменения содержания антоцианов при засухе, которые также были выявлены у Quercus virginiana, Quercus geminata, Quercus fusiformis и Quercus oleoides в проведенном ранее исследовании (Ramírez-Valiente et al., 2015).
Содержание флавонолов в контрольной группе снижалось, что может свидетельствовать о меньшем эффекте стресса на сеянцы Q. robur. Однако при отсутствии полива содержание флавонолов повышалась к поздним срокам эксперимента. Это согласуется с исследованием (Rivas-Ubach et al., 2014), где было показано, что при засухе возрастает содержание флавонолов в листьях Quercus ilex.
Отсутствие изменения транскрипционной активности DREB2A, rbcL и AAO при поливе может свидетельствовать о том, что растения не испытывали стресс. В тоже время в условиях засухи мы обнаружили резкое повышение экспрессии транскрипционного фактора DREB2A с его последующим снижением к концу второй недели эксперимента, в то время как активность rbcL и AAO постепенно повышалась. Возможно, DREB2A запускает быстрый ответ на засуху (Nakashima et al., 2000; Soma et al., 2021), поэтому в последующем его активность снижается (Qiu et al., 2024). Исходя из этого, мы предполагаем, что в среднесрочной перспективе начинают преобладать другие адаптационные механизмы. Это предположение согласуется с тем, что под воздействием длительной засухи начинают работать ABA-зависимые механизмы (Sreenivasulu et al., 2012; Zhao et al., 2017), обеспечивающие приспособление растений к стрессовым условиям. Возможно, так как ABA-зависимые механизмы являются энергозатратными и повышается активность генов фотосинтетической системы, в частности, согласно нашему исследованию – rbcL, что подтверждается результатами корреляционного анализа. При этом известно, что транскрипционная активность rbcL напрямую связана с интенсивностью цикла Кальвина (Paul 1998, Suzuki et al., 2001), которая возможно увеличивается при стрессе.
Заключение
В результате изучения ответной реакции сеянцев Q. robur на засуху в моделируемых условиях были выявлены изменения на молекулярно-генетическом уровне. У Q. robur при засухе в листьях увеличивалось содержание флавонолов и антоцианов, обладающих антиоксидантными свойствами. Кроме того, было зафиксировано, что в зависимости от длительности воздействия стресс-фактора – засухи изменяется транскрипционная активность генов. Наибольший уровень транскрипционной активности DREB2A приходился на первую неделю эксперимента, а на второй снижался до уровня контрольной группы. В то же время была выявлены обратная тенденция для AAO, где через две недели эксперимента транскрипционная активность достигла максимального значения. При этом была выявлена сильная корреляция между уровнями экспрессии AAO и rbcL и умеренная у DREB2A и rbcL, а также AAO и DREB2A в условиях засухи на второй неделе. Это свидетельствует о том, что AAO и DREB2A играют важную роль в механизмах адаптации к засухе у Q. robur. Полученные данные способствуют расширению знаний о возможных механизмах адаптации к засухе у древесных растений. В прикладных исследованиях эти знания могут быть использованы для создания улучшенных генотипов Q. robur путем классической гибридизации или генной инженерии, включая геномное редактирование.
Благодарности
Мы выражаем благодарность лаборатории Биоэкологии древесных растений ФГБНУ «Федеральный научный центр агроэкологии, комплексных мелиораций и защитного лесоразведения Российской академии наук» за предоставление прибора Dualex Scientific+™.
Финансирование
Работа выполнена в рамках государственного задания Министерства науки и высшего образования Российской Федерации № FNFE-2025-0013 «Управление экспрессией генов, вовлеченных в процесс адаптации к неблагоприятным факторам внешней среды, для улучшения хозяйственно-ценных признаков древесно-кустарниковых и культурных растений, используемых в агролесомелиорации».
Литература / References
- Andersson I., Backlund A. Structure and function of Rubisco // Plant Physiology and Biochemistry. 2008. Vol. 46. № 3. P. 275-291. https://doi.org/10.1016/j.plaphy.2008.01.001
- Cai L. J., Ji X. Y., Hou M. F., Hao Y. F., Wu Q., Song L. L. The influence of ABA on the photosynthesis of the rare and endangered Emmenopterys henryi under salt stress // Biol Plantarum. 2023. Vol. 67. P. 1-8. https://doi.org/10.32615/bp.2022.036
- Carmo-Silva E., Sharwood R. E. Rubisco and its regulation – major advances to improve carbon assimilation and productivity // Journal of Experimental Botany. 2023. Vol. 74. № 2. P. 507-509. https://doi.org/10.1093/jxb/erac475
- Carter G. A., Knapp A. K. Leaf optical properties in higher plants: linking spectral characteristics to stress and chlorophyll concentration // American journal of botany. 2001. Vol. 88. № 4. P. 677-684. https://doi.org/10.2307/2657068
- Chen K., Li G. J., Bressan R. A., Song C. P., Zhu J. K., Zhao Y. Abscisic acid dynamics, signaling, and functions in plants // Journal of integrative plant biology. 2020. Vol. 62. № 1. P. 25-54. https://doi.org/10.1111/jipb.12899
- Chen W., Miao Y., Ayyaz A., Hannan F., Huan, Q., Ulhassan Z., et al. Purple stem Brassica napus exhibits higher photosynthetic efficiency, antioxidant potential and anthocyanin biosynthesis related genes expression against drought stress // Frontiers in Plant Science. 2022. Vol. 13. P. 936696. https://doi.org/10.3389/fpls.2022.936696
- Diethelm A. C., Reichelt M., Dilts T. E., Farlin J. P., Marlar A., Pringle E. G. Climatic history, constraints, and the plasticity of phytochemical traits under water stress // Ecosphere. 2022. Vol. 13. № 8. P. e4167. https://doi.org/10.5281/zenodo.6518931
- Egawa C., Kobayashi F., Ishibashi M., Nakamura T., Nakamura C., Takumi S. Differential regulation of transcript accumulation and alternative splicing of a DREB2 homolog under abiotic stress conditions in common wheat // Genes & genetic systems. 2006. Vol. 81. № 2. P. 77-91. https://doi.org/10.1266/ggs.81.77
- Eldridge D. J., Ding J., Dorrough J., Delgado-Baquerizo M., Sala O., Gross N., et al. Hotspots of biogeochemical activity linked to aridity and plant traits across global drylands // Nature Plants. 2024. Vol. 10. № 5. P. 760-770. https://doi.org/10.1038/s41477-024-01670-7
- Engelbrecht B. M. J., Kursar T. A. Comparative drought-resistance of seedlings of 28 species of co-occurring tropical woody plants // Oecologia. 2003. Vol. 136. № 3. P. 383-393. https://doi.org/10.1007/s00442-003-1290-8
- Feeley K. J., Bravo-Avila C., Fadrique B., Perez T. M., Zuleta D. Climate-driven changes in the composition of New World plant communities // Nature Climate Change. 2020. Vol. 10. № 10. P. 965-970. https://doi.org/10.1038/s41558-020-0873-2
- Fujita Y., Fujita M., Shinozaki K., Yamaguchi-Shinozaki K. ABA-mediated transcriptional regulation in response to osmotic stress in plants // Journal of plant research. 2011. Vol. 124. № 4. P. 509-525. https://doi.org/10.1007/s10265-011-0412-3
- Galka M. M., Rajagopalan N., Buhrow L. M., Nelson K. M., Switala J., Cutler A. J., et al. Identification of interactions between abscisic acid and ribulose-1, 5-bisphosphate carboxylase/oxygenase // PLoS One. 2015. Vol. 10. № 7. P. e0133033. https://doi.org/10.1371/journal.pone.0133033
- Gallé A., Haldimann P., Feller U. Photosynthetic performance and water relations in young pubescent oak (Quercus pubescens) trees during drought stress and recovery // New Phytologist. 2007. Vol. 174. № 4. P. 799-810. https://doi.org/10.1111/j.1469-8137.2007.02047.x
- González E. M. Drought stress tolerance in plants // International journal of molecular sciences. 2023. Vol. 24. № 7. P. 6562. https://doi.org/10.3390/ijms24076562
- Gould K. S., Neill S. O., Vogelmann T. C. A unified explanation for anthocyanins in leaves? // Advances in Botanical Research. 2002. Vol. 37. P. 167-192. https://doi.org/10.1016/S0065-2296(02)37049-6
- Grjibovski A. M., Ivanov S. V., Gorbatova M. A. Analysis of quantitative data in two non-independent groups using statistica and SPSS software: parametric and non-parametric tests // Science and Healthcare. 2016. Vol. 5. P. 5-29. https://doi.org/10.34689/SH.2016.18.5.001
- Hermans K., McLeman R. Climate change, drought, land degradation and migration: exploring the linkages // Current opinion in environmental sustainability. 2021. Vol. 50. P. 236-244. https://doi.org/10.1016/j.cosust.2021.04.013
- Hrmova M., Hussain S. S. Plant transcription factors involved in drought and associated stresses // International journal of molecular sciences. 2021. Vol. 22. № 11. P. 5662. https://doi.org/10.3390/ijms22115662
- Ikegami K., Okamoto M., Seo M., Koshiba T. Activation of abscisic acid biosynthesis in the leaves of Arabidopsis thaliana in response to water deficit // Journal of plant research. 2009. Vol. 122. № 2. P. 235-243. https://doi.org/10.1007/s10265-008-0201-9
- Jafarnia S., Akbarinia M., Hosseinpour B., Modarres Sanavi S. A. M., Salami S. A. Effect of drought stress on some growth, morphological, physiological, and biochemical parameters of two different populations of Quercus brantii // iForest-Biogeosciences and Forestry. 2018. Vol. 11. № 2. P. 212. https://doi.org/10.3832/ifor2496-010
- Jan R., Asif S., Asaf S., Lubna, Khan Z., Kim K. M. Unveiling the protective role of anthocyanin in rice: insights into drought-induced oxidative stress and metabolic regulation // Frontiers in Plant Science. 2024. Vol. 15. P. 1397817. https://doi.org/10.3389/fpls.2024.1397817
- Javaid M. M., Florentine S., Mahmood A., Wasaya A., Javed T., Sattar A., et al. Interactive effect of elevated CO2 and drought on physiological traits of Datura stramonium // Frontiers in Plant Science. 2022. Vol. 13. P. 929378. https://doi.org/10.3389/fpls.2022.929378
- Kim J. S., Mizoi J., Yoshida T., Fujita Y., Nakajima J., Ohori T., et al. An ABRE promoter sequence is involved in osmotic stress-responsive expression of the DREB2A gene, which encodes a transcription factor regulating drought-inducible genes in Arabidopsis // Plant and Cell Physiology. 2011. Vol. 52. № 12. P. 2136-2146. https://doi.org/10.1093/pcp/pcr143
- Kulik K. N., Belyaev A. I., Pugacheva A. M. The role of protective afforestation in drought and desertification control in agro-landscapes // Arid Ecosystems. 2023. Vol. 13. № 1. P. 1-10. https://doi.org/10.1134/S2079096123010079
- Kurganova I., Lopes de Gerenyu V., Khoroshaev D., Myakshina T., Sapronov D., Zhmurin V. Temperature sensitivity of soil respiration in two temperate forest ecosystems: the synthesis of a 24-year continuous observation // Forests. 2022. Vol. 13. № 9. P. 1374. https://doi.org/10.3390/f13091374
- Lata C., Prasad M. Role of DREBs in regulation of abiotic stress responses in plants // Journal of experimental botany. 2011. Vol. 62. № 14. P. 4731-4748. https://doi.org/10.1093/jxb/err210
- Livak K. J., Schmittgen T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method // methods. 2001. Vol. 25. № 4. P. 402-408. https://doi.org/10.1006/meth.2001.1262
- Manna M., Thakur T., Chirom O., Mandlik R., Deshmukh R., Salvi P. Transcription factors as key molecular target to strengthen the drought stress tolerance in plants // Physiologia Plantarum. 2021. Vol. 172. № 2. P. 847-868. https://doi.org/10.1111/ppl.13268
- Mikhin V. I., Mikhina E. A. Formation of English oak protective plants in the Central Chernozem region of Russia // Forestry engineering journal. 2018. Vol. 4. P. 109-117. https://doi.org/10.12737/article_5c1a321965cf38.69751554
- Nakashima K., Shinwari Z. K., Sakuma Y., Seki M., Miura S., Shinozaki K., Yamaguchi-Shinozaki K. Organization and expression of two Arabidopsis DREB2 genes encoding DRE-binding proteins involved in dehydration-and high-salinity-responsive gene expression // Plant molecular biology. 2000. Vol. 42. № 4. P. 657-665. https://doi.org/10.1023/A:1006321900483
- Nikolaeva M. K., Maevskaya S. N., Shugaev A. G., Bukhov N. G. Effect of drought on chlorophyll content and antioxidant enzyme activities in leaves of three wheat cultivars varying in productivity // Russian Journal of Plant Physiology. 2010. Vol. 57. № 1. P. 87-95. https://doi.org/10.1134/S1021443710010127
- Paul J. H. Carbon cycling: molecular regulation of photosynthetic carbon fixation // Microbial ecology. 1996. Vol. 32. № 3. P. 231-245. https://doi.org/10.1007/BF00183060
- Qiao M., Hong C., Jiao Y., Hou S., Gao H. Impacts of drought on photosynthesis in major food crops and the related mechanisms of plant responses to drought // Plants. 2024. Vol. 13. № 13. P. 1808. https://doi.org/10.3390/plants13131808
- Qiu W., Li H., Song Y., Ding J., Chen S., Ma C., Yu B. Genome-wide analysis of DREB genes in sugar beet and their potential functions in response to drought stress // Sugar Tech. 2024. Vol. 26. № 5. P. 1306-1322. https://doi.org/10.1007/s12355-023-01337-1
- Ramírez-Valiente J. A., Koehler K., Cavender-Bares J. Climatic origins predict variation in photoprotective leaf pigments in response to drought and low temperatures in live oaks (Quercus series Virentes) // Tree Physiology. 2015. Vol. 35. № 5. P. 521-534. https://doi.org/10.1093/treephys/tpv032
- Rini D. W. I. S. Sequence variation of DREB2 gene as a potential molecular marker for identifying resistant plants toward drought stress // Nusantara Bioscience. 2019. Vol. 11. № 1. P. 35-43. https://doi.org/10.13057/nusbiosci/n110107
- Rivas-Ubach A., Gargallo-Garriga A., Sardans J., Oravec M., Mateu-Castell L., Pérez-Trujillo M., et al. Drought enhances folivory by shifting foliar metabolomes in Quercus ilex trees // New Phytologist. 2014. Vol. 202. № 3. P. 874-885. https://doi.org/10.1111/nph.12687
- Sabouri H., Pezeshkian Z., Taliei F., Akbari M., Kazerani B. Detection of closely linked QTLs and candidate genes controlling germination indices in response to drought and salinity stresses in barley // Scientific Reports. 2024. Vol. 14. № 1. P. 15656. https://doi.org/10.1038/s41598-024-66452-9
- Semerikova S. A., Aliev K. U., Semerikov N. V., Semerikov V. L. Phylogeography of oak species in the Caucasus based on results of chloroplast DNA analysis // Russian Journal of Genetics. 2023. Vol. 59. № 7. P. 669-684. https://doi.org/10.1134/S1022795423070104
- Shi P, Wang Y, Yin C, Fan K, Qian Y, Chen G. Mitigating saturation effects in rice nitrogen estimation using Dualex measurements and machine learning // Front Plant Sci. 2024. № 15. P. 1518272. https://doi.org/10.3389/fpls.2024.1518272.
- Shi X., Tian Q., Deng P., Zhan, W., Jing W. The rice aldehyde oxidase OsAO3 gene regulates plant growth, grain yield, and drought tolerance by participating in ABA biosynthesis // Biochemical and biophysical research communications. 2021. Vol. 548. P. 189-195. https://doi.org/10.1016/j.bbrc.2021.02.047
- Singh K., Chandra A. DREBs-potential transcription factors involve in combating abiotic stress tolerance in plants // Biologia. 2021. Vol. 76. № 10. P. 3043-3055. https://doi.org/10.1007/s11756-021-00840-8
- Soma F., Takahashi F., Yamaguchi-Shinozaki K., Shinozaki K. Cellular phosphorylation signaling and gene expression in drought stress responses: ABA-dependent and ABA-independent regulatory systems // Plants. 2021. Vol. 10. № 4. P. 756. https://doi.org/10.3390/plants10040756
- Sreenivasulu N., Harshavardhan V. T., Govind G., Seiler C., Kohli A. Contrapuntal role of ABA: does it mediate stress tolerance or plant growth retardation under long-term drought stress? // Gene. 2012. Vol. 506. № 2. P. 265-273. https://doi.org/10.1016/j.gene.2012.06.076
- Suzuki Y., Makino A., Mae T. Changes in the turnover of Rubisco and levels of mRNAs of rbcL and rbcS in rice leaves from emergence to senescence // Plant, Cell & Environment. 2001. Vol. 24. № 12. P. 1353-1360. https://doi.org/10.1046/j.0016-8025.2001.00789.x
- Trivedi D. K., Gill S. S., Tuteja N. Abscisic acid (ABA): biosynthesis, regulation, and role in abiotic stress tolerance // Abiotic stress response in plants. 2016. P. 315-326. https://doi.org/10.3390/plants13111451
- Yan X., Cheng P., Zhang Q., Li X., He J., Yan X., et al. Comparisons of climate change characteristics in typical arid regions of the Northern Hemisphere // Frontiers in Environmental Science. 2022. Vol. 10. P. 1033326. https://doi.org/10.3389/fenvs.2022.1033326
- Zamolodchikov D. G., Kaganov V. V., Mostovaya A. S. The Effect of Forest Plantations on Carbon Dioxide Emission from Soils in the Volga and Don Regions // Russian Journal of Ecology. 2023. Vol. 54. № 6. P. 584-593. https://doi.org/10.1134/S1067413623070160
- Zhao L., Cai Z., Li Y., Zhang Y. Engineering Rubisco to enhance CO2 utilization // Synthetic and Systems Biotechnology. 2024. Vol. 9. № 1. P. 55-68. https://doi.org/10.1016/j.synbio.2023.12.006
- Zhao Y., Gao J., Im Kim J., Chen K., Bressan R. A., Zhu J. K. Control of plant water use by ABA induction of senescence and dormancy: an overlooked lesson from evolution // Plant and Cell Physiology. 2017. Vol. 58. № 8. P. 1319-1327. https://doi.org/10.1093/pcp/pcx086
- Zybinskaya P. A., Tretyakova A. V. Selection of reference genes for quantitative gene expression analyses in pedunculate oak (Quercus robur L.) using real-time quantitative PCR // E3S Web of Conferences. – EDP Sciences, 2024. Vol. 548. P. 07007. https://doi.org/10.1051/e3sconf/202454807007
Статья поступила в редакцию 9 февраля 2026 г.
Поступила после доработки 20 февраля 2026 г.
Принята к печати 13 марта 2026 г.
Received 9, February, 2026
Revised 20, February, 2026
Accepted 13, March, 2026