Введение:
Белки семейства WNT играют ключевую роль в регуляции эмбриогенеза, тканевых взаимодействий и репродуктивных процессов (Гребенникова и др., 2016; Есенеева и др., 2017). Среди них гены WNT1 и WNT3A участвуют в каскадах сигнальных путей, необходимых для формирования нервных тканей, скелета и осевого развития. WNT1 регулирует морфогенез центральной нервной системы, включая развитие среднего мозга и мозжечка (McMahon, Bradley, 1990; Ikeya M, e.a., 1997; Lee e.a., 2000; Костенко и др., 2018). Его недостаточность приводит к тяжелым порокам, таким как гипоплазия мозга и деформации черепа. Кроме того, WNT1 влияет на репродуктивную систему, нарушая дифференцировку половых желез и фертильность (Татарский, 2016; Alset, e.a., 2024; Ebrahimi, e.a., 2026). WNT3A обеспечивает сегментирование сомитов и осевое развитие организма. Нарушения его экспрессии вызывают отклонения в опорно-двигательной системе и наследственные заболевания (Joeng e.a.,2017; Снимщикова и др., 2024). Продукты этих генов также влияют на репродукцию: недостаток WNT1 снижает фертильность, а нарушения WNT3A негативно сказываются на беременности. Генетические изменения в локусах WNT1 и WNT3A ассоциированы с проблемами зачатия и вынашивания. Глубокие знания о молекулярных особенностях этих генов имеют клиническое значение для лечения бесплодия и профилактики нарушений развития. Цель работы — сравнить нуклеотидные последовательности генов WNT1 и WNT3A у человека и мыши, определить уровень гомологии и исследовать связь результатов с особенностями эмбриогенеза и репродуктивной функцией.
Материалы и методы:
Для сравнительного анализа использовались гены семейства Wnt: WNT1 и WNT3A человека (Homo sapiens) и мыши (Mus musculus). Координаты генов определялись на основе аннотаций геномов hg38 (человек) и mm39 (мышь) из Ensembl (release 109). Геномные последовательности и аннотации были загружены из открытых источников: Все последовательности были приведены к формату FASTA, а аннотации — к формату GTF. Последовательности каждого гена человека и мыши были разделены по отдельным файлам FASTA, а затем объединены для проведения множественного выравнивания, извлечение последовательностей генов из соответствующих геномов проводили с помощью bedtools getfasta. Сравнительный анализ нуклеотидных последовательностей генов WNT1 и WNT3A человека (Homo sapiens, сборка hg38) и мыши (Mus musculus, сборка mm39) проводился на полных геномных регионах, включая кодирующие и некодирующие области (промоторы, интроны, UTR). Координаты регионов:
WNT1: человек — chr12:48,978,322–48,982,620 (4299 п.н.);
мышь — chr15:98,687,738–98,691,711 (3974 п.н.).
WNT3A: человек — chr1:228,006,998–228,061,271 (54 274 п.н.);
мышь — chr11:59,138,859–59,181,579 (42 721 п.н.).
Выравнивание последовательностей человека и мыши проводилось с использованием MAFFT (v7.490). Вычислялись следующие параметры: Общая длина выравнивания, количество совпадающих нуклеотидов, процент идентичности, количество гепов. Для оценки локальной консервативности применялась скользящая оконная стратегия с окном 100 нуклеотидов и шагом 50 нуклеотидов. Результаты сохранялись в таблицах CSV и визуализировались графически. Для каждой последовательности рассчитывалось количество нуклеотидов A, T, G и C, а также GC-состав. Данные визуализировались в виде столбчатых диаграмм для наглядного сравнения между человеком и мышью. Консервативные мотивы длиной ≥6 нуклеотидов идентифицировались по позициям совпадений в выравнивании. Для каждого мотива вычислялись координаты в геномах человека и мыши. Для мотивов человека была выполнена аннотация относительно геномных элементов с использованием PyRanges и GTF-аннотаций. Мотивы классифицировались по типу: экзон, интрон, промотор или межгенное пространство. Анализ проводился с использованием следующих инструментов: BEDTools v2.30 для извлечения последовательностей; MAFFT v7.490 для множественного выравнивания; Python 3.11 с библиотеками: BioPython, pandas, numpy, matplotlib, PyRanges. Все скрипты были автоматизированы в единый пайплайн для воспроизводимого анализа.
Результаты исследований:
Множественное выравнивание последовательностей с помощью MAFFT выявило существенные различия в степени консервативности между генами (табл. 1). Для гена WNT1 общая длина выравнивания составила 4409 п.н., с 2990 совпадающими позициями из 3862 сравниваемых (исключая гэпы), что соответствует 77.42% идентичности. Количество гэпов — 547, что указывает на относительно компактную структуру гена с умеренными вставками/делециями. Для гена WNT3A результаты значительно отличаются: длина выравнивания — 59 158 п.н., совпадения — 19 398 из 37 835 сравниваемых позиций (51.27% идентичности), с высоким числом гэпов (21 323). Это отражает более низкую эволюционну.ю консервативность. Разница в идентичности (77.42% для гена WNT1 vs. 51.27% для гена WNT3A) подчеркивает более строгую консервативность гена WNT1, вероятно, из-за его роли в критических процессах деятельность центральной нервной системы, в то время как ген WNT3A демонстрирует большую видоспецифичную вариабельность в регуляторных элементах.
Таблица 1. Результаты выравнивания гена WNT1, WNT3A у человека и мыши
| Ген | Длина выравнивания | Совпадения | Выровненные позиции | Пропуски/гепы | Идентичность в % |
|---|---|---|---|---|---|
| WNT1 | 4409 | 2990 | 3862 | 547 | 77.421 |
| WNT3A | 59158 | 19398 | 37835 | 21323 | 51.27 |
Далее нами был проведен анализ нуклеотидного состава последовательностей генов WNT1 и WNT3A у человека и мыши. Для гена WNT1 (рис. 1) наблюдается заметное обогащение нуклеотидами G и C: содержание C достигает примерно 30–33%, G — 27–28%, что приводит к повышенному GC-составу (около 60.7% у человека и 57.5% у мыши). Содержание A и T ниже (A ~19–20%, T ~20–22%). Такая GC-богатость типична для генов, вовлеченных в регуляцию развития нервной системы, и может быть связана с наличием CpG-островков в промоторных регионах, способствующих эпигенетической регуляции. Для гена WNT3A (рис. 1) распределение нуклеотидов более сбалансированное: A и T составляют около 24–25% каждый, C и G — 24–27%, с общим GC-составом около 51.9% у человека и 50.0% у мыши. Небольшие различия между видами (человек слегка GC-богаче) могут отражать видоспецифичные особенности интронных последовательностей в этом протяженном гене.
В обоих генах нуклеотидный состав у человека и мыши сходен, что указывает на сохранение базовых физико-химических свойств ДНК (например, стабильности и изгибания), важных для регуляторных взаимодействий.
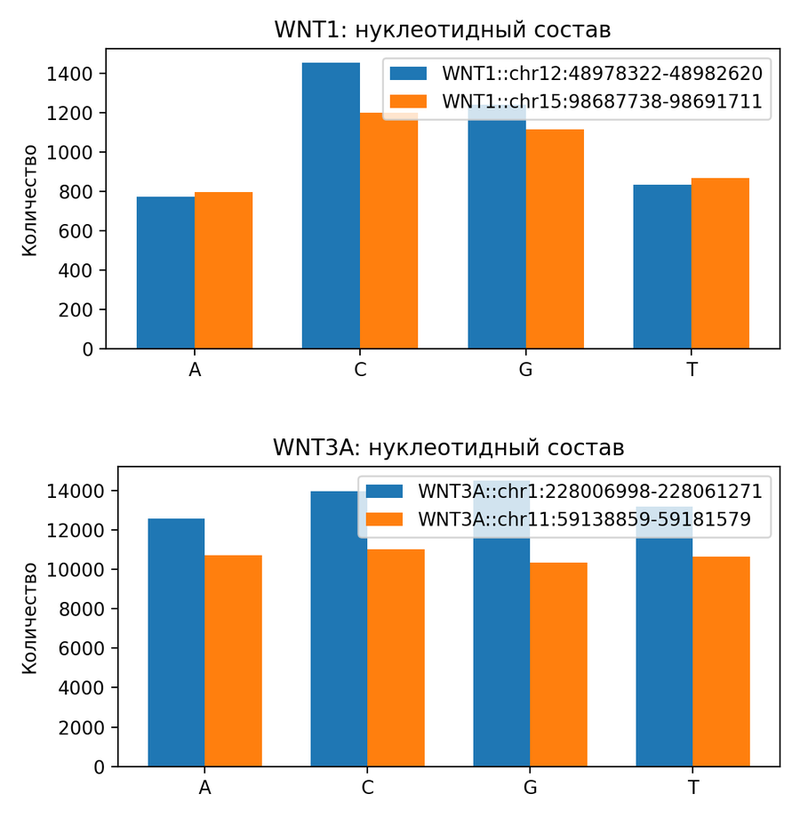
Рисунок 1. Сравнение нуклеотидных составов гена WNT1 и WNT3A у человека и мыши.
*Синим отмечен нуклеотидный состав человека, оранжевым –мыши.
Распределение GC-состава вдоль выравнивания оценивалось в скользящем окне100 п.н., шаг 50 п.н.) отдельно для последовательностей человека (синяя линия) и мыши (оранжевая линия).

Рисунок 2. Динамика GC-состава вдоль выравнивания гена WNT1 и WNT3A у человека и мыши.
Значения плотности распределения CpG-островков у человека и мыши колеблются преимущественно в диапазоне 40–70%, с несколькими выраженными пиками до 70–80% (особенно в 5'-области). Совпадение пиков GC с областями высокой консервативности указывает на возможное наличие CpG-островков в промоторных и экзонных регионах, играющих роль в эпигенетической регуляции экспрессии гена.
Плотность распределения CpG-островков в гене WNT3A у человека и мыши более вариабельна по сравнению с геном WNT1. Кривые человека и мыши тесно переплетаются, с частыми острыми пиками до 80–100% и провалами до 20%. Общий уровень GC-состава ниже (в среднем 45–55%). Высокие пики, как правило, соответствуют консервативным блокам, что подтверждает обогащение GC в функционально значимых участках (экзоны, регуляторные элементы), тогда как протяженные интронные области характеризуются сниженным GC-составом. Сходство траекторий GC-кривых между видами наиболее выражено в консервативных регионах, что отражает эволюционное давление не только на первичную последовательность, но и на физико-химические свойства ДНК (стабильность, изгибание), важные для взаимодействия с регуляторными факторами. В таблице 2 представлен GC-состав консервативных мотивов гена. Для WNT1/Wnt1 средний GC-состав мотивов практически идентичен фоновому уровню региона, с минимальной отрицательной разницей (–0.34%). Это указывает на то, что консервативные мотивы не обогащены или обеднены GC-нуклеотидами относительно всего локуса и отражают общую высокую GC-насыщенность гена WNT1/Wnt1.
Таблица 2. GC-состав консервативных мотивов гена по сравнению с фоновым
| Ген | GC-фон региона (%) | Средний GC мотивов (%) | ΔGC (мотивы – фон) (%) |
|---|---|---|---|
| WNT1 | 52.51 | 54.41 | +1.90 |
| WNT3A | 62.66 | 62.31 | -0.34 |
В отличие от гена WNT1, для гена WNT3A наблюдается заметное положительное отклонение: средний GC-состав консервативных мотивов на 1.90% выше фонового. Это свидетельствует о слабом, но статистически значимом обогащении GC-нуклеотидами в строго консервативных участках. Такое обогащение часто ассоциировано с функционально важными регуляторными элементами (CpG-островки, сайты связывания транскрипционных факторов), что особенно актуально для протяженного гена WNT3A с большим количеством интронных регуляторных последовательностей. Полученные различия подчёркивают разные стратегии эволюционной консервативности: в гене WNT1/Wnt1 сохраняется весь GC-богатый локус в целом, тогда как в гене WNT3A селективное давление направлено на GC-обогащённые функциональные «островки».
При выравнивании последовательностей человека и мыши для гена WNT1 идентифицировано множество строго консервативных мотивов (непрерывные участки полной идентичности длиной ≥6 п.н.). Ниже приведены самые длинные мотивы от 68 ло 20 нуклеотидов по убыванию ( таблица 3).
Таблица 3. Самые протяженные консервативные мотивы гена WNT1/Wnt1
| Начало | Конец | Длина (н.п.) | sequence | GC% | Кол-во CpG -островков |
|---|---|---|---|---|---|
| 3328 | 3396 | 68 | GAGCGCTGCAACTGCACCTTCCACTGGTGCTGCCACGTCAGCTGCCGCAACTGCACGCACACGCGCGT | 66.2 | 5 |
| 1411 | 1460 | 49 | GGGCCCCACCTCTTCGGCAAGATCGTCAACCGAGGTGGGTGCCCAGGAA | 65.3 | 5 |
| 3160 | 3207 | 47 | CACGACCTCGTCTACTTCGAGAAATCGCCCAACTTCTGCACGTACAG | 53.2 | 7 |
| 2342 | 2377 | 35 | GAGTTCGTGGACTCCGGGGAGAAGGGGCGGGACCT | 68.6 | 3 |
| 2378 | 2413 | 35 | CGCTTCCTCATGAACCTTCACAACAACGAGGCAGG | 54.3 | 3 |
| 2276 | 2308 | 32 | GGGGGCCCCGACTGGCACTGGGGGGGCTGCAG | 81.3 | 4 |
| 404 | 435 | 31 | CCGCTCTGCCCGCAGCCCTGGCTGCCAACAG | 74.2 | 1 |
| 334 | 364 | 30 | GCCAGGCAGGCCATGGGGCTCTGGGCGCTG | 76.7 | 1 |
| 4313 | 4343 | 30 | ATCAGTATTTCCTTCCACTGTAGCTATTAG | 36.7 | 0 |
| 3034 | 3063 | 29 | GTGCTGCGCGACCGCTTCGACGGCGCCTC | 75.9 | 2 |
| 3100 | 3129 | 29 | TCGCGGGCGGAGCTGCTGCGCCTGGAGCC | 79.3 | 3 |
| 3298 | 3327 | 29 | GGCCACCGCACGCGCACGCAGCGCGTCAC | 79.3 | 4 |
| 4151 | 4180 | 29 | TGTAAATAAAACTATTTATTGTGCTGGGT | 27.6 | 0 |
| 4071 | 4098 | 27 | ACCCCTTCCTGTCCTGCCTCCTCATCA | 59.3 | 0 |
| 1312 | 1338 | 26 | CCGGGGATCCTGCACAGCGTGAGTGG | 69.2 | 3 |
| 2998 | 3024 | 26 | TGGATGCGGCTGCCCACGCTGCGCGC | 76.9 | 5 |
| 436 | 461 | 25 | AGTGGCCGATGGTGGTAAGTGAGCT | 56.0 | 2 |
| 1255 | 1276 | 21 | CTCGAGCCCAGTCTGCAGCTG | 66.7 | 2 |
| 4025 | 4046 | 21 | GGAGCCATTGAACAGCTGTGA | 52.4 | 1 |
| 1205 | 1225 | 20 | TAGCCTCCTCCACGAACCTG | 60.0 | 1 |
| 2183 | 2203 | 20 | ATCACCTCCGCCGGGGTCAC | 70.0 | 2 |
| 2255 | 2275 | 20 | GACTACCGGCGGCGCGGCCC | 85.0 | 3 |
| 2959 | 2979 | 20 | TGCCACGGGATGTCCGGCTC | 70.0 | 5 |
| 3561 | 3581 | 20 | CCTCCTACCTGGGGACTCCT | 65.0 | 2 |
Как видно из таблицы 3 многие консервативные мотивы обладают высоким GC-составом (до 85%), что характерно для потенциальных регуляторных элементов (CpG-островки). Большинство длинных мотивов расположены в регионах некодирующей ДНК (интроны, промотор), что указывает на возможную регуляторную функцию. В представленном наборе не выявлено известных сайтов связывания транскрипционных факторов, однако высокое содержание CpG (до 7 в одном мотиве) предполагает потенциал для эпигенетической регуляции. Средний GC длинных мотивов (>20 п.н.) составляет ≈65–70%, что выше среднего по геному человека и согласуется с ранее рассчитанным небольшим отрицательным ΔGC (–0.34%) — консервативные участки отражают общий GC-богатый фон локуса гена WNT1. Эти строго консервативные последовательности могут представлять функционально значимые элементы (энхансеры, сайленсеры, сайты сплайсинга), сохранённые эволюцией несмотря на разделение видов ~75 млн лет назад.
В таблице 4 приведены результаты выравнивания последовательностей гена WNT3A/Wnt3a человека и мыши. Как видно из представленных результатов для гена WNT3A/Wnt3a обнаружено значительно меньшее количество консервативных мотивов (≥6 п.н.), чем для гена WNT1, при этом практически все они короткие. В таблице 4 приведены примеры наиболее длинных мотивов из доступного набора данных (представлены мотивы длиной ≥10 п.н.; самый длинный в выборке — 15 п.н.).
Таблица 4. Наиболее протяженные консервативные мотивы гена WNT3A/Wnt3a
| start | end | length | sequence | GC% | CpG_count | |
|---|---|---|---|---|---|---|
| 52644 | 52659 | 15 | ATATATATATATATA | 0.0 | 0 | |
| 6264 | 6276 | 12 | AGACAGGGTCTC | 58.3 | 0 | |
| 2091 | 2101 | 10 | CACCCTCGGG | 80.0 | 0 | |
| 11933 | 11943 | 10 | TTTTTGAGAC | 30.0 | 0 | |
| 12179 | 12189 | 10 | GATTACAGGC | 50.0 | 0 | |
| 27552 | 27562 | 10 | GTTCTCTCTG | 50.0 | 0 | |
| 35381 | 35391 | 10 | CTGAGCTTGG | 60.0 | 0 |
Самый длинный консервативный мотив в гене WNT3A/Wnt3a человека и мыши содержит 15 п.н., представляет собой повторяющийся поли-A/T (ATATATATATATATA), с GC = 0%. Такие A/T-богатые последовательности часто встречаются в интронных и регуляторных регионах и могут участвовать в формировании структур ДНК или быть частью микро-/минисателлитов. Среди консервативных мотивов длиной 10–12 п.н. GC-состав варьирует от 0% до 80%. Все представленные мотивы расположены в регионах в интронах и не содержат аннотированных сайтов связывания транскрипционных факторов (TFBS = none) или CpG (0 в выборке). Низкое содержание CpG в длинных мотивах согласуется с общим умеренным GC-фоном гена WNT3A/Wnt3a (≈52.5%), но ранее рассчитанное положительное ΔGC (+1.90%) указывает, что в целом по всем мотивам (включая короткие) наблюдается слабое обогащение GC в консервативных участках. В отличие от гена WNT1, где длинные мотивы часто GC-богаты и потенциально регуляторны, в гене WNT3A/Wnt3a даже относительно длинные консервативные блоки включают A/T-богатые повторы, что может отражать видоспецифичные структурные особенности интронов или эволюционно сохранённые элементы, связанные с регуляцией на уровне хроматина или сплайсинга.
Обсуждение:
Проведённый сравнительный анализ геномных последовательностей WNT1/Wnt1 и WNT3A/Wnt3a выявил различия в общей нуклеотидной идентичности и пространственном распределении консервативных участков. Однако эти различия следует интерпретировать прежде всего с учётом неодинаковой архитектуры исследуемых локусов. По современной RefSeq-аннотации человеческий и мышиный локусы WNT1/Wnt1 занимают приблизительно 4,3 и 4,0 кб соответственно, тогда как WNT3A/Wnt3a имеют значительно большую протяжённость — около 54,3 кб у человека и 42,7 кб у мыши. Оба гена состоят из четырёх экзонов, поэтому различия в длине локусов преимущественно обусловлены интронными последовательностями. Общая идентичность 77,42% для WNT1/Wnt1 и 51,27% для WNT3A/Wnt3a характеризует не только функционально значимые последовательности, но и различия в длине интронов, содержании повторов и количестве вставок и делеций. Несмотря на различия в общей идентичности геномных локусов, кодируемые ими белки высококонсервативны: WNT1 человека имеет приблизительно 98% аминокислотной идентичности с мышиным Wnt1, а WNT3A — около 96% с мышиным Wnt3a. Раннее прямое сравнение человеческого и мышиного WNT1 показало наличие только четырёх аминокислотных замен в белке длиной 370 аминокислот (van Ooyen e.a., 1985). Оба белка находятся под сильным отбором, а пониженная идентичность локуса WNT3A/Wnt3a преимущественно отражает ускоренную эволюцию некодирующей ДНК, а не снижение консервативности основной биохимической функции WNT3A. Высокий GC-состав (≈62%) в регионе и мотивах (ΔGC ≈ –0.34%) указывает на наличие CpG-островков в промоторных областях, что может способствовать эпигенетической регуляции экспрессии, включая метилирование ДНК, влияющее на развитие мозга и репродуктивные нарушения. В отличие от гена WNT1/Wnt1, для гена WNT3A/Wnt3a идентичность между геном человека и мыши ниже (51.27%), с высокой гетерогенностью в скользящем окне (частые провалы <30%) и большим числом коротких мотивов, преимущественно в интронах (61.2%). Это отражает большую пластичность регуляторных элементов в протяжённом гене, ответственном за сегментацию сомитов и осевое развитие. Положительное ΔGC в мотивах (+1.90%) предполагает селективное обогащение GC в функционально значимых «островках», таких как энхансеры, что согласуется с ролью WNT3A в мезодермальной индукции и органогенезе. Низкая консервативность некодирующих областей может объяснять видоспецифичные адаптации, включая влияние на фертильность и беременность. Сравнение нуклеотидного состава показывает, что оба гена имеют схожий баланс A/T/G/C между видами, но ген WNT1 более GC-богат (58–60% ), чем ген WNT3A/Wnt3a (50–52%), что типично для генов с строгой экспрессионной регуляцией в нервной ткани. Консервативные мотивы в гене WNT1/Wnt1 часто длинные и GC-насыщенные (до 85%), потенциально содержащие сайты для транскрипционных факторов, в то время как в гене WNT3A преобладают короткие A/T-богатые повторы, которые могут влиять на стабильность РНК или хроматиновую организацию. Более низкая общая идентичность локуса WNT3A/Wnt3a согласуется с его большей протяжённостью и значительной долей интронной ДНК. Ранее было показано, что экзонно-интронная организация кластера WNT3A–WNT9A в целом сохраняется у человека и мыши, тогда как межгенное расстояние и состав некодирующих последовательностей существенно различаются
Полученные данные подтверждают ключевую роль этих генов в эмбриогенезе и репродуктивных процессах, обосновывая использование мышиной модели для изучения патологий, связанных с WNT-сигнализацией. Результаты могут способствовать разработке новых подходов к диагностике и терапии бесплодия, врожденных аномалий и связанных заболеваний, создавая основу для дальнейших молекулярно-генетических исследований.
Полученные результаты показывают, что различия между локусами WNT1/Wnt1 и WNT3A/Wnt3a проявляются главным образом на уровне протяжённости и эволюционной изменчивости некодирующей ДНК, тогда как кодируемые WNT-белки высококонсервативны. Компактный локус WNT1/Wnt1 содержит протяжённые консервативные блоки, часть которых может соответствовать известным элементам регуляции экспрессии в развивающейся нервной системе. Более протяжённый и интронно-богатый локус WNT3A/Wnt3a характеризуется меньшей глобальной идентичностью, что согласуется с эволюционным оборотом некодирующих последовательностей при сохранении основной функции WNT3A в формировании задней оси и параксиальной мезодермы. Выявленные мотивы следует рассматривать как кандидатные регуляторные элементы, требующие сопоставления с эпигеномными данными и экспериментальной проверки. Результаты обосновывают применение мышиной модели для изучения консервативных белковых и морфогенетических функций WNT1 и WNT3A, но не предполагают полной взаимозаменяемости регуляторных последовательностей человека и мыши.
Работа выполнена при финансовой поддержке Министерства науки и высшего образования РФ в рамках государственного задания в сфере научной деятельности FENW-2026-0030.
Список литературы:
- Гребенникова Т. А., Белая Ж. Е., Рожинская Л. Я., Мельниченко Г. А. Канонический сигнальный путь Wnt/β-катенин: от истории открытия до клинического применения // Терапевтический архив. — 2016. — Т. 88, № 10. — С. 74–81. — DOI: 10.17116/terarkh201688674-81.
- Есенеева Ф. М., Шалаев О. Н., Хамошина М. Б., Оразмурадов А. А., Киселев В. И., Салимова Л. Я. Активация WNT-сигнального пути у пациенток с миомой матки // Исследования и практика в медицине. — 2017. — Т. 4, № S2. — С. 51.
- Костенко Н. К., Юнси Г. А., Василенко С. А., Абдурахимов У. Э. Значение Wnt-сигнализации в эмбрио- и онтогенезе // Крымский журнал экспериментальной и клинической медицины. — 2018. — № 2. — С. —.
- Снимщикова И. А., Плотникова М. О., Кулакова А. С., Якушкина Н. Ю. WNT-сигнальный путь и его связь с метаболическими нарушениями: роль DVL-1 и WIF-1 // Российский иммунологический журнал. — 2024. — Т. 27, № 3. — С. 545–552.
- Татарский В. В. Сигнальный путь Wnt: перспективы фармакологического регулирования // Успехи молекулярной онкологии. — 2016. — Т. 3, № 1. — С. 28–31.
- Alset D. et al. Genetic variations of Wnt/β-catenin signaling pathway microRNA regulators as novel prenatal biomarkers of fetal growth restriction syndrome //Gene Reports. – 2024. – Т. 35. – С. 101914.
- Ebrahimi, A., Heidary, Z., & Zaki-Dizaji, M. (2026). PPP1R1C: a specific placental mRNA biomarker for distinguishing preeclampsia from healthy pregnancies // BMC Pregnancy and Childbirth.
- Ikeya M, Lee SM, Johnson JE, McMahon AP, Takada S. Wnt signalling required for expansion of neural crest and CNS progenitors // Nature. 1997;389:966–970. URL: https://doi.org/10.1038/40146
- Joeng KS, Lee YC, Lim J, Chen Y, Jiang MM, Munivez E, et al. Osteocyte-specific WNT1 regulates osteoblast function during bone homeostasis // J Clin Invest. 2017;127(7):2678–2688. URL: https://doi.org/10.1172/JCI92617
- Lee SMK, Tole S, Grove E, McMahon AP. A local Wnt-3a signal is required for development of the mammalian hippocampus. Development. 2000;127(3):457–467. URL: https://doi.org/10.1242/dev.127.3.457
- McMahon AP, Bradley A. The Wnt-1 proto-oncogene is required for development of a large region of the mouse brain // Cell. 1990;62(6):1073–1085. URL: https://doi.org/10.1016/0092-8674(90)90385-R
- van Ooyen A, Kwee V, Nusse R. The nucleotide sequence of the human int-1 mammary oncogene; evolutionary conservation of coding and non-coding sequences. EMBO J. 1985;4(11):2905–2909
Статья поступила в редакцию 3 марта 2026 г.
Поступила после доработки 12 марта 2026 г.
Принята к печати 20 марта 2026 г.
Received 3, March, 2026
Revised 12, March, 2026
Accepted 20, March, 2026