Введение
Пандемия коронавирусной инфекции COVID-19, вызванной вирусом SARS-CoV-2, стала одной из наиболее значимых глобальных проблем здравоохранения последних лет. Несмотря на значительный прогресс в понимании патогенеза заболевания, механизмы, определяющие индивидуальную вариабельность клинических проявлений инфекции, остаются предметом активного изучения. Накопленные данные свидетельствуют о том, что наряду с возрастом, полом и сопутствующими заболеваниями существенное влияние на тяжесть COVID-19 оказывают генетические особенности организма хозяина.
Ключевую роль в ограничении репликации SARS-CoV-2 играют механизмы врожденного иммунитета, прежде всего интерфероновые сигнальные пути (Zhou et al.,2020). Интерфероны I и III типов обеспечивают ранний противовирусный ответ, индуцируя экспрессию генов с противовирусной активностью (Platanias, 2005). Нарушения функционирования интерфероновой системы рассматриваются в качестве одного из факторов развития тяжелых форм COVID-19. Одним из важных компонентов данного сигнального каскада является тирозинкиназа TYK2, принадлежащая к семейству Janus-киназ и участвующая в передаче сигналов от рецепторов интерферонов и ряда провоспалительных цитокинов (Platanias, 2005). Генетические варианты локуса TYK2 ассоциированы с иммуновоспалительными и аутоиммунными заболеваниями, включая ревматоидный артрит, системную красную волчанку и рассеянный склероз (Stahl et al., 2010; Sawcer et al., 2011; Bentham et al., 2015). Кроме того, JAK/STAT-сигнальный путь, в функционировании которого участвует TYK2, рассматривается как ключевая мишень патогенетической терапии COVID-19 (Kalil et al., 2021; RECOVERY Collaborative Group, 2022).
Помимо белок-кодирующих генов, важную роль в регуляции иммунного ответа играют микроРНК — небольшие некодирующие молекулы РНК, контролирующие экспрессию генов на посттранскрипционном уровне. Среди них особое внимание привлекают miR-146a и miR-155, участвующие в поддержании баланса между активацией противовирусной защиты и ограничением избыточного воспаления (Taganov et al., 2006; O’Connell et al., 2012).
МикроРНК miR-146a является одним из ключевых отрицательных регуляторов врожденного иммунитета. Полиморфизм rs2910164 гена miR-146a способен влиять на процесс созревания микроРНК и уровень ее экспрессии (Jazdzewski et al., 2008). В последние годы показано, что снижение уровня miR-146a может быть связано с неблагоприятным течением COVID-19 и выраженным воспалительным ответом (Sabbatinelli et al., 2021).
В отличие от miR-146a, микроРНК miR-155 преимущественно рассматривается как положительный регулятор иммунной активации. Она участвует в контроле сигнальных путей NF-κB и JAK/STAT, влияет на продукцию провоспалительных цитокинов и функциональную активность различных популяций иммунокомпетентных клеток (O’Connell et al., 2012). Изменения экспрессии miR-155 были выявлены при различных вирусных инфекциях, а также у пациентов с COVID-19, что позволяет рассматривать данную микроРНК как один из потенциальных молекулярных маркеров тяжести заболевания (Taha et al., 2025; Kassif-Lerner et al., 2022).
Следует отметить, что развитие заболевания COVID-19 представляет собой сложный многофакторный процесс, в котором клинический эффект может определяться не только независимым влиянием отдельных локусов, но и их взаимодействием.
Учитывая ключевую роль TYK2, miR-146a и miR-155 в регуляции противовирусного и воспалительного ответа, представляется перспективным изучение как индивидуального вклада соответствующих генетических вариантов, так и их возможных эпистатических взаимодействий в формировании тяжести течения COVID-19.
Цель настоящего исследования оценить ассоциации генетических вариантов TYK2 rs11085727, miR-146a rs2910164 и miR-155 rs767649 с тяжестью течения COVID-19, а также изучить возможные межгенные взаимодействия между указанными локусами.
Материал и методы
Объектами исследования послужили образцы крови больных COVID-19. Рекрутинг пациентов осуществлялся в период с 12.03.2021 г по 15.01.2022 г. Исследование проводилось в постморбидный период (не менее 2х месяцев после заболевания). Для участи исследовании в медицинский центр «Наука» (Ростов-на-Дону) обратился 451 пациент. Критерием включения в исследование было наличие антител к спайковому (S) белку SARS-CoV-2, иммуноглобулинов G. Для проведения дальнейшего исследования было отобрано 217 пациентов. В соответствии с рекомендациями Министерства здравоохранения Российской Федерации (Временные методические рекомендации - профилактика, диагностика и лечение новой коронавирусной инфекции (COVID-19)) пациенты были разделена на 2 группы в зависимости от тяжести симптомов (113 - с легкой и 104 -с тяжелой формой COVID-19). Участникам проводились анкетирование для получения анамнестических данных: дата заболевания, возраст, пол, данные компьютерной томографии, наличие сопутствующих заболеваний. При осмотре оценивались следующие показатели: артериальное давление (АД), уровень насыщения крови кислородом (SpO2), частота сердечных сокращений (ЧСС), рост и вес. Информированное согласие было получено от всех участников исследования.
Клинико-демографическая характеристика групп пациентов с легкой и тяжелой степенью COVID-19 представлена в таблице 1.
Таблица 1. Клинико-демографическая характеристика групп пациентов.
| Показатель | Лёгкое течение (n=113) | Тяжёлое течение (n=104) | p-value |
|---|---|---|---|
| Пол, n (%) | Пол, n (%) | Пол, n (%) | Пол, n (%) |
| Мужчины | 41 (36,3%) | 33 (31,7%) | 0.46 |
| Женщины | 72 (63,7%) | 71 (68,3%) | |
| Возраст, годы (M ± SD) | 46,0 ± 15,1 | 55,1 ± 11,1 | <0.001 |
| ИМТ, кг/м² (M ± SD) | 25,6 ± 5,0 | 29,2 ± 5,1 | <0.001 |
Данные представлены как среднее арифметическое ± стандартное отклонение (M ± SD) для количественных переменных и абсолютное число (n) с процентной долей (%) для категориальных переменных.
Молекулярно-генетические методы
Молекулярно-генетические исследования проводились в лаборатории биологии развития и организации генома на кафедре генетики в Академии биологии и медицины им. Д.И. Ивановского Южного федерального университета (Ростов-на-Дону).
Для оценки вклада генетического варианта тирозинкиназы 2 в предрасположенность к тяжёлому течению COVID-19 был проведён анализ полиморфизма гена TYK2 rs11085727 (C>T) методом оценки длины рестрикционных фрагментов. При этом использовали праймеры (прямой 5'-TGAATAACCACCACACTCCT-3', обратный 5'- CACTGCGCCTGACCTATT-3') и рестриктзазу Alu1.
Для оценки вклада генетических вариантов генов микроРНК, участвующих в регуляции воспалительного ответа, в предрасположенность к тяжёлому течению COVID-19 был проведён анализ двух полиморфных локусов: miR-146a rs2910164 (G>C) и miR-155 rs767649 (T>A). Генотипирование выполняли методом аллель-специфической ПЦР. Идентификацию продуктов амплификации проводили методом электрофореза либо в режиме детекции в реальном времени.
Статистическая обработка данных выполнена с использованием программного пакета IBM SPSS Statistics 27.0 (IBM Corp., Armonk, NY, USA). Сравнение групп проводилось с помощью U-критерия Манна–Уитни (возраст, ИМТ) и критерия χ² Пирсона (генотипы, пол). При ожидаемой частоте менее 5 применяли точный критерий Фишера. Равновесие Харди–Вайнберга (HWE) рассчитывали с использованием веб-инструмента SNPStats (https://www.snpstats.net/start.htm) (Solé et al., 2006). Для изучения межгенных взаимодействий использовали алгоритм многомерного снижения размерности (Multifactor Dimensionality Reduction - MDR) версии 3.0.2 (He et al., 2009).
Результаты и обсуждение
Перед анализом ассоциаций была проведена оценка соответствия распределения генотипов равновесию Харди–Вайнберга (HWE) в каждой группе отдельно (таблица 2). Для всех исследованных генетических локусов распределение генотипов соответствовало HWE (p> 0,05 для всех сравнений).
Таблица 2. Равновесие Харди–Вайнберга в исследуемых группах
| Локус | Группа | χ² | p-value |
|---|---|---|---|
| miRNA-146a (rs2910164) | Лёгкие | 1,16 | 0,281 |
| Тяжёлые | 1,39 | 0,239 | |
| miR-155 (rs767649) | Лёгкие | 0,10 | 0,757 |
| Тяжёлые | 0,01 | 0,920 | |
| TYK2 rs11085727 | Лёгкие | 0,437 | 0,508 |
| Тяжёлые | 1,349 | 0,245 |
При анализе полиморфного локуса rs11085727 гена TYK2 статистически значимых различий между группами лёгкого и тяжёлого течения COVID-19 не обнаружено (χ² = 2,98, p = 0,225 для генотипов; χ² = 3,10, p = 0,078 для аллелей). Распределение генотипов и аллелей представлены в таблице 3.
Таблица 3. Распределение генотипов и аллелей полиморфизма TYK2 rs11085727 у пациентов с лёгким и тяжёлым течением COVID-19
| Генотипы/Аллели | Пациенты с лёгкой формой COVID-19 (n=113) | Пациенты с тяжёлой формой COVID-19 (n=104) | χ² | p-value | OR (95% ДИ) |
|---|---|---|---|---|---|
| Генотипы | Генотипы | Генотипы | Генотипы | Генотипы | Генотипы |
| CC | 52 (46,0%) | 60 (57,7%) | 2,98 | 0,225 | 1,60 (0,93–2,75) |
| CT | 47 (41,6%) | 35 (33,7%) | 2,98 | 0,225 | 0,71 (0,41–1,23) |
| TT | 14 (12,4%) | 9 (8,7%) | 2,98 | 0,225 | 0,67 (0,28–1,62) |
| CT+TT | 99 (87,6%) | 95 (91,3%) | 2,97 | 0,085 | 0,62 (0,37–1,06) |
| CC+CT | 99 (87,6%) | 95 (91,3%) | 0,67 | 0,413 | 1,49 (0,62–3,58) |
| Аллели | Аллели | Аллели | Аллели | Аллели | Аллели |
| Аллель C | 151 (66,8%) | 155 (74,5%) | 3,10 | 0,078 | 1,46 (0,96–2,21) |
| Аллель T | 75 (33,2%) | 53 (25,5%) | 3,10 | 0,078 | 0,69 (0,45–1,04) |
Генотип CC в группе лёгкого течения составил 46,0%, в группе тяжёлого - 57,7% (OR = 1,60; 95% ДИ 0,93–2,75); генотип TT - 12,4% и 8,7% соответственно (OR = 0,67; 95% ДИ 0,28–1,62). Частота минорного аллеля T в группе лёгких составила 33,2%, в группе тяжёлых - 25,5% (OR = 0,69; 95% ДИ 0,45-1,04), что может указывать на потенциальную связь аллеля T со снижением риска тяжёлого течения COVID-19.Аллель C встречался в 74,5% случаев в группе тяжёлого течения и 66,8% случаев в группе легкого течения заболевания. Несмотря на отсутствие статистической значимости, выявленные тенденции позволяют предположить возможное участие данного локуса в формировании индивидуальной восприимчивости к тяжёлому течению COVID-19, что требует проверки на более расширенных выборках.
Ген TYK2 кодирует нерецепторную тирозинкиназу, которая является одним из четырех членов семейства Janus-киназ (O'Shea et al., 2012). TYK2 играет критическую роль в передаче сигналов от рецепторов цитокинов, таких как интерфероны I и III типа, интерлейкины IL-6, IL-10, IL-12 и IL-23, посредством JAK-STAT сигнального пути (Schindler et al., 2007; Ghoreschi et al., 2009). Эти цитокины являются центральными регуляторами врожденного и адаптивного иммунитета, а также воспалительных процессов. Исследуемый полиморфизм rs11085727 представляет собой однонуклеотидный вариант, расположенный в интронной области гена TYK2 (согласно данным dbSNP, NCBI). Интронные варианты, как правило, не приводят к изменению аминокислотной последовательности белка, но могут влиять на стабильность мРНК, регуляцию экспрессии гена или быть в сильном неравновесном сцеплении с другими функциональными вариантами (Ward et al., 2009). Несмотря на интронную локализацию, полиморфизм rs11085727 представляет интерес в связи с тем, что ген TYK2 неоднократно идентифицировался в полногеномных ассоциативных исследованиях как один из ключевых генов предрасположенности к аутоиммунным и воспалительным заболеваниям, включая рассеянный склероз, псориаз, воспалительные заболевания кишечника, системную красную волчанку, сахарный диабет 1 типа и ревматоидный артрит (Stahl et al., 2010; Bradfield et al., 2011; Sawcer et al., 2011; Liu et al., 2015; Bentham et al., 2015). Эти данные свидетельствуют о важной роли TYK2 в регуляции иммунного ответа и поддержании баланса между противовирусной защитой и воспалением. Вместе с тем функциональное значение rs11085727 остается недостаточно изученным, а его вклад в восприимчивость к тяжелому течению COVID-19 пока не получил однозначного подтверждения.
Особый интерес к TYK2 в контексте COVID-19 обусловлен его центральной ролью в сигнальных путях интерферонов I и III типа, обеспечивающих ранний противовирусный ответ организма (Platanias, 2005; Park et al., 2020). Нарушение интерферонового ответа рассматривается как один из ключевых механизмов развития тяжелых форм SARS-CoV-2-инфекции. Тогда как гиперпродукция провоспалительных цитокинов способствует формированию системного воспаления и повреждению тканей (Del Valle et al., 2020). Клиническая эффективность ингибиторов JAK-STAT-сигналинга, включая барицитиниб, дополнительно подтверждает значимость данного молекулярного каскада в патогенезе COVID-19 (Kalil et al., 2021; RECOVERY Collaborative Group, 2022).
Для оценки вклада генетических вариантов генов микроРНК, участвующих в регуляции воспалительного ответа, в предрасположенность к тяжёлому течению COVID-19 был проведён анализ двух полиморфных локусов: miRNA-146a rs2910164 (G>C) и miR-155 rs767649 (T>A). Распределение генотипов и аллелей в группах лёгкого и тяжёлого течения представлено в таблице 4.
Таблица 4. Распределение частот генотипов и аллелей полиморфных вариантов rs2910164 miRNA-146a и rs767649 miRNA-155 у пациентов с COVID-19
| Пациенты с легкой формой COVID-19 (n=113) | Пациенты с тяжелой формой COVID-19 (n=104) | χ² | p-value | OR (95% ДИ) | |
|---|---|---|---|---|---|
| miRNA-146a rs2910164 (C> G) | miRNA-146a rs2910164 (C> G) | miRNA-146a rs2910164 (C> G) | miRNA-146a rs2910164 (C> G) | miRNA-146a rs2910164 (C> G) | miRNA-146a rs2910164 (C> G) |
| Генотипы | Генотипы | Генотипы | Генотипы | Генотипы | Генотипы |
| CC | 29 (25,7%) | 22 (21,2%) | 0,78 | 0,674 | 0,78 (0,41–1,47) |
| CG | 62 (54,9%) | 58 (55,8%) | 0,78 | 0,674 | 1,04 (0,61–1,76) |
| GG | 22 (19,5%) | 24 (23,1%) | 0,78 | 0,674 | 1,24 (0,65–2,38) |
| CG+GG | 84 (74,3%) | 82 (78,8%) | 0,62 | 0,430 | 1,29 (0,69–2,41) |
| CC+CG | 91 (80,5%) | 80 (76,9%) | 0,43 | 0,511 | 0,80 (0,41–1,55) |
| Аллели | Аллели | Аллели | Аллели | Аллели | Аллели |
| Аллель C | 120 (53,1%) | 102 (49,0%) | 0,01 | 0,920 | 0,85 (0,59–1,22) |
| Аллель G | 106 (46,9%) | 106 (51,0%) | 0,01 | 0,920 | 1,18 (0,82–1,70) |
| miRNA-155 rs767649 ( T>A) | miRNA-155 rs767649 ( T>A) | miRNA-155 rs767649 ( T>A) | miRNA-155 rs767649 ( T>A) | miRNA-155 rs767649 ( T>A) | miRNA-155 rs767649 ( T>A) |
| Генотипы | Генотипы | Генотипы | Генотипы | Генотипы | Генотипы |
| TT | 46 (40,7%) | 41 (39,4%) | 0,06 | 0,970 | 0,95 (0,55–1,63) |
| TA | 51 (45,1%) | 49 (47,1%) | 0,06 | 0,970 | 1,08( 0,63–1,86) |
| AA | 16 (14,2%) | 14 (13,5%) | 0,06 | 0,970 | 0,94 (0,44–2,03) |
| TA+AA | 67 (59,3%) | 63 (60,6%) | 0,04 | 0,846 | 1,06 (0,62–1,81) |
| TT+TA | 97 (85,8%) | 90 (86,5%) | 0,02 | 0,884 | 1,06(0,48–2,34) |
| Аллели | Аллели | Аллели | Аллели | Аллели | Аллели |
| Аллель T | 143 (63,3%) | 131 (63,0%) | 0,11 | 0,740 | 0,99(0,68–1,43) |
| Аллель A | 83 (36,7%) | 77 (37,0%) | 0,11 | 0,740 | 1,01 (0,70–1,47) |
При анализе полиморфного локуса rs2910164 (G>C) гена miR-146a статистически значимых различий между группами лёгкого и тяжёлого течения COVID-19 не обнаружено (χ² = 0,79, p = 0,674 для генотипов; χ² = 0,01, p = 0,920 для аллелей). Генотип CC в группе лёгкого течения составил 25,7%, в группе тяжёлого - 21,2% (OR = 0,78; 95% ДИ 0,41-1,47); генотип GG - 19,5% и 23,1% соответственно (OR = 1,24; 95% ДИ 0,65–2,38). Частота минорного аллеля C в группе лёгких составила 53,1%, в группе тяжёлых - 49,0% (OR = 0,85; 95% ДИ 0,59-1,22). Полученные результаты свидетельствуют об отсутствии ассоциации исследуемого полиморфизма с тяжестью течения COVID-19 в исследованных группах.
Полиморфизм rs 2910164 miR-146a расположен в последовательности предшественника микроРНК-146a и регулирует экспрессию генов-мишеней, участвующих в контроле воспалительного ответа (Ngo et al.,2025). Было показано, что данный вариант способен влиять на созревание микроРНК и уровень экспрессии зрелой miR-146a, что может отражаться на регуляции генов врождённого иммунитета (Jazdzewski et al., 2008).
MiR-146a рассматривается как один из основных отрицательных регуляторов воспалительного ответа, ограничивающий активацию сигнального пути NF-κB (Nuclear Factor kappa B) посредством подавления экспрессии IRAK1 (Interleukin-1 Receptor-Associated Kinase 1) и TRAF6 (TNF Receptor Associated Factor 6) (Taganov et al., 2006). Благодаря этому miR-146a играет важную роль в поддержании баланса между эффективной противоинфекционной защитой и чрезмерным воспалением.
Особое значение miR-146a приобретает в контексте COVID-19, поскольку тяжёлое течение заболевания характеризуется выраженным нарушением иммунного ответа и гиперпродукцией провоспалительных цитокинов (Del Valle et al., 2020). В ряде исследований было показано, что снижение экспрессии miR-146a ассоциируется с усилением воспалительных процессов и неблагоприятным течением SARS-CoV-2-инфекции (Sabbatinelli et al., 2021; Gaytán-Pacheco et al., 2022). Однако полученные нами результаты свидетельствуют о том, что генетическая вариабельность rs2910164 сама по себе не оказывает значимого влияния на риск развития тяжёлой формы COVID-19. Вероятно, изменения уровня miR-146a при коронавирусной инфекции в большей степени определяются особенностями воспалительного процесса.
МикроРНК miR-155 является одним из ключевых регуляторов иммунного ответа и участвует в активации макрофагов, дифференцировке Т- и В-лимфоцитов, а также контроле продукции цитокинов (O'Connell et al., 2012).
Клиническая значимость miR-155 в патогенезе COVID-19 была продемонстрирована в исследовании Kassif-Lerner и соавторов (2022), где было отмечено прогрессирующее снижение уровня этой микроРНК по мере утяжеления состояния пациентов. Так, концентрация циркулирующей miR-155 у пациентов с тяжелой формой заболевания была в 5 раз ниже, чем в контрольной группе, в то время как при легкой форме это снижение было менее выраженным (в 2,5 раза). Авторы подчеркивают, что miR-155 обладает высокой прогностической ценностью в отношении смертности от COVID-19, и мониторинг её уровня при поступлении в стационар может стать важным инструментом в клинической практике (Kassif-Lerner et al., 2022)
Полиморфизм rs767649 гена miR-155 расположен в промоторной области и, по данным некоторых исследований, может влиять на транскрипционную активность гена (Zheng et al., 2018).
Анализ распределения генотипов rs767649 (T>A) гена miR-155 в исследуемой выборке не выявил статистически значимых различий между группами лёгкого и тяжёлого течения COVID-19 ((χ² = 0,06, p = 0,970 для генотипов; χ² = 0,11, p = 0,740 для аллелей).). Генотип TT в группе лёгкого течения составил 40,7%, в группе тяжёлого - 39,4% (OR = 0,95; 95% ДИ 0,55–1,63); генотип AA - 14,2% и 13,5% соответственно (OR = 0,94; 95% ДИ 0,44-2,03). Частота минорного аллеля A в группе лёгких составила 36,7%, в группе тяжёлых - 37,0% (OR = 1,01; 95% ДИ 0,70-1,47).
Несмотря на отсутствие ассоциации rs767649 с тяжестью COVID-19 в нашем исследовании, нельзя исключить, что вклад miR-155 в патогенез COVID-19 определяется преимущественно изменениями уровня её экспрессии в ходе инфекции, тогда как влияние отдельных генетических вариантов регуляторной области miR-155 может быть недостаточным для формирования статистически значимого эффекта на клиническое течение заболевания.
Для выявления генетических факторов риска тяжелого течения COVID-19 был также проведен анализ межгенных взаимодействий с использованием метода Multifactor Dimensionality Reduction (MDR). Исследовали взаимодействие трех полиморфных локусов: rs11085727 гена TYK2, rs2910164 гена miR-146a и rs767649 гена miR-155.
Для более детального изучения вклада каждого локуса и характера их парных взаимодействий был построен график Фрюхтермана-Рейнгольда (Рисунок 1). Этот метод позволяет количественно оценить независимый эффект каждого полиморфизма (энтропию) и тип межгенного взаимодействия (синергизм или антагонизм).
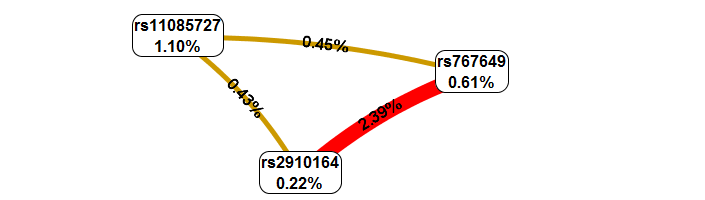
Рисунок 1. График Фрюхтермана-Рейнгольда с типами взаимодействий между полиморфизмами. Каждый блок полиморфизма содержит значение энтропии (%), что отражает его независимый эффект. Цвета и значения между полями указывают эффекты взаимодействия. Положительные значения представляют собой синергетический эффект, а отрицательные — антагонизм. Цвет линии указывает тип межгенного взаимодействия. Красный цвет обозначает высокую степень синергичного взаимодействия, оранжевый — меньшую степень синергичного взаимодействия; коричневый — промежуточный этап между совместными действиями и антагонизмом (отсутствие связи или независимость эффектов отдельных локусов); зеленый и синий — антагонизм эффектов с меньшей и большей степенью, соответственно.
Анализ графика Фрюхтермана–Рейнгольда показал, что наибольший независимый вклад в информационную структуру модели вносил локус rs11085727 гена TYK2 (энтропия 1,10%), тогда как для rs767649 гена miR-155 и rs2910164 гена miR-146a значения энтропии составили 0,61% и 0,22% соответственно. Несмотря на относительно низкие значения индивидуального вклада, между исследованными локусами были выявлены признаки положительного взаимодействия. Наиболее выраженное взаимодействие наблюдалось между rs2910164 и rs767649, для которого величина информационной энтропии составила 2,39%. Полученные данные могут свидетельствовать о том, что влияние указанных локусов на течение заболевания реализуется не только посредством независимых эффектов, но и через их совместное участие в регуляции иммунного ответа.
Анализ отдельных локусов (1-факторные модели) показал, что ни один из исследованных локусов сам по себе не обладает достаточной предсказательной силой для определения тяжести COVID-19. Максимальная взвешенная точность для однофакторных моделей составила 0.561 (для rs11085727), что лишь незначительно превышает случайное предсказание (0.5). Это указывает на то, что индивидуальные эффекты этих полиморфизмов, вероятно, малы или отсутствуют в данной популяции.
Вместе с тем при анализе комбинаций локусов наблюдалось последовательное увеличение точности моделей. Наиболее информативной двухлокусной моделью оказалась комбинация rs2910164 miR146a и rs767649 miR-155, для которой взвешенная точность достигала 0,602.
Оптимальной моделью, согласно результатам MDR, стала трехлокусная модель, включающая все три исследованных локуса: rs11085727 гена TYK2, rs2910164 miR-146a, rs767649 гена miR-155. Эта модель продемонстрировала наивысшую взвешенную точность -0.676 (таблица 5).
Таблица 5 Модель межгенных взаимодействий полиморфных локусов генов TYK2 rs11085727, miR-146a rs2910164 и miR-155 rs776746 у пациентов с легкой и тяжелой формой COVID-19
| Комбинации локусов в модели | Контрольная взвешенная точность | Опытная взвешенная точность | p значение (χ2) | Воспроизводимость модели (CV consistency) |
|---|---|---|---|---|
| TYK2 rs 11085727 miR-146a rs2910164 miR-155 rs767649 | 0,67 | 0,63 | <0,0001* (24,3) | 10/10 |
Наиболее сильная ассоциация с тяжёлым течением COVID-19 выявлена для комбинации генотипов TYK2 rs11085727 CC, miR-146a rs2910164 GG, miR-155 rs767649 TT (OR = 2,24; 95% ДИ 1,00–5,03; χ² = 4,05; p = 0,044). Протективные комбинации (CC TYK2 rs11085727-CG miR-146a rs2910164 - TT miR-155 rs767649 и CT TYK2 rs11085727 - GG miR-146a rs2910164 -TT miR-155 rs767649) показали тенденцию к защитному эффекту, однако не достигли статистической значимости (p> 0,05).
Выявленные эпистатические взаимодействия указывают на то, что вклад отдельных генетических вариантов в риск тяжелого течения COVID-19 реализуется не изолированно, а модулируется аллельным составом других генов. Представляется важным, что выявленная модель объединяет локусы, относящиеся к различным уровням иммунной регуляции. Современные представления о патогенезе COVID-19 свидетельствуют о том, что тяжесть заболевания определяется не только эффективностью противовирусной защиты, но и способностью организма контролировать интенсивность воспалительного ответа (Del Valle et al., 2020; Park et al.,2020). В связи с этим, выявленные взаимодействия между локусами могут указывать на важность их совместного участия в регуляции иммунного ответа, определяющего особенности клинического течения COVID-19
Особый интерес представляет выявленная комбинация генотипов TYK2 rs11085727 CC, miR-146a rs2910164 GG и miR-155 rs767649 TT, ассоциированная с повышенным риском тяжелого течения COVID-19. Ген TYK2 участвует в передаче сигналов интерферонов и других цитокинов, обеспечивающих раннюю противовирусную защиту, тогда как miR-146a и miR-155 являются ключевыми регуляторами воспалительных реакций и врождённого иммунитета (Platanias, 2005; Taganov et al., 2006; O’Connell et al., 2012). Предполагаем, что комбинация (CC, GG, TT) создаёт провоспалительный дисбаланс: минорный аллель C rs2910164 снижает продукцию зрелой miR-146a (ослабляя торможение NF-κB), а генотип TT rs767649 повышает экспрессию miR-155 (усиливая активацию макрофагов). Совместно с генотипом CC rs11085727, ассоциированным с повышенной активностью TYK2, это может способствовать гиперпродукции IL-6, TNF-α и CXCL10 — ключевых медиаторов «цитокинового шторма» при тяжёлом COVID-19 (Del Valle et al., 2020). В условиях SARS-CoV-2 изменение такого регуляторного баланса может способствовать развитию гипервоспалительного ответа и цитокин-опосредованного повреждения тканей, характерных для тяжёлых форм заболевания (Del Valle et al., 2020; Sabbatinelli et al., 2021).
Заключение
В настоящем исследовании проведена оценка возможной роли полиморфных вариантов rs11085727 гена TYK2, rs2910164 гена miR146a и rs767649 гена miR155 в формировании предрасположенности к тяжелому течению COVID-19. Анализ распределения генотипов и аллелей не выявил статистически значимых ассоциаций отдельных исследованных полиморфных локусов с тяжестью заболевания. Вместе с тем для полиморфизма TYK2 rs11085727 отмечена тенденция к ассоциации минорного аллеля T со снижением риска тяжёлого течения COVID-19 (OR = 0,69; 95% ДИ: 0,45–1,04; p = 0,078), что может свидетельствовать о его потенциальном участии в формировании индивидуальных особенностей иммунного ответа.
Проведенный MDR-анализ позволил выявить эпистатические взаимодействия между исследованными локусами. Наиболее информативной оказалась трехлокусная модель, включавшая гены TYK2 rs11085727, miR146A rs2910164 и miR155 rs767649. Модель характеризовалась сбалансированной точностью 0,675 при 10-кратной кросс-валидации. Особый интерес представляет выявленная комбинация генотипов TYK2 rs11085727 CC, miR146a rs2910164 GG и miR155 rs767649 TT, ассоциированная с повышенным риском тяжелого течения COVID-19 ((OR = 2,24; 95% ДИ 1,00–5,03; χ² = 4,05; p = 0,044). Полученные данные свидетельствуют о том, что влияние генетических факторов на клинические проявления SARS-CoV-2-инфекции может определяться не только отдельными полиморфными вариантами, но и особенностями их совместного функционирования в регуляции иммунного ответа.
Таким образом, результаты исследования указывают на возможную роль эпистатических взаимодействий между генами TYK2, miR146a и miR155 в формировании тяжести течения COVID-19. Полученные данные расширяют представления о генетических механизмах индивидуальной восприимчивости к тяжелым формам заболевания и обосновывают целесообразность дальнейшего изучения межгенных взаимодействий в патогенезе коронавирусной инфекции на более крупных выборках.
Работа выполнена при финансовой поддержке Министерства науки и высшего образования РФ в рамках государственного задания в сфере научной деятельности FENW-2026-0030.
Список литературы:
- Временные методические рекомендации МЗ РФ «Профилактика, диагностика и лечение новой коронавирусной инфекции – COVID-19» (версия № 18 от 26.10.2023 г.) / МЗ РФ // Москва, 2023. – 250 с.
- Bentham J., et al. Genetic association analyses implicate aberrant regulation of innate and adaptive immunity genes in the pathogenesis of systemic lupus erythematosus. Nature Genetics. 2015;47(12):1457-1464. doi:10.1038/ng.3434
- Bradfield J.P., et al. A genome-wide meta-analysis of six type 1 diabetes cohorts identifies multiple associated loci. PLoS Genetics. 2011;7(9):e1002293. doi:10.1371/journal.pgen.1002293
- Del Valle D.M., et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nature Medicine. 2020;26(10):1636-1643. doi:10.1038/s41591-020-1051-9
- Gaytán-Pacheco N., Ibáñez-Salazar A., Herrera-Van Oostdam A.S., et al. miR-146a, miR-221, and miR-155 are Involved in Inflammatory Immune Response in Severe COVID-19 Patients. Diagnostics. 2023;13(1):133. doi:10.3390/diagnostics13010133
- He, H., Oetting, W.S., Brott, M.J. et al. Power of multifactor dimensionality reduction and penalized logistic regression for detecting gene-gene Interaction in a case-control study. BMC Med Genet 10, 127 (2009). https://doi.org/10.1186/1471-2350-10-127
- Jazdzewski K., et al. Common SNP in pre-miR-146a decreases mature miR expression and predisposes to papillary thyroid carcinoma. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(20):7269-7274. doi:10.1073/pnas.0802682105
- Kalil A.C., et al. Baricitinib plus Remdesivir for Hospitalized Adults with Covid-19. New England Journal of Medicine. 2021;384(9):795-807. doi:10.1056/NEJMoa2031994
- Kassif-Lerner R., et al. miR-155: A Potential Biomarker for Predicting Mortality in COVID-19 Patients. Journal of Personalized Medicine. 2022;12(2):324. doi:10.3390/jpm12020324
- Liu J.Z., et al. Association analyses identify 38 susceptibility loci for inflammatory bowel disease and highlight shared genetic risk across populations. Nature Genetics. 2015;47(9):979-986. doi:10.1038/ng.3359
- 11.Ngo T.H., Ho C.Q., Tran S.K. Genetic association of microRNA-146a polymorphisms with the severity of coronary artery lesions in acute myocardial infarction. PLoS One. 2025;20(12):e0339345. doi:10.1371/journal.pone.0339345
- O’Connell R.M., et al. microRNA regulation of inflammatory responses. Annual Review of Immunology. 2012;30:295-312. doi:10.1146/annurev-immunol-020711-075013
- Park A., Iwasaki A. Type I and Type III Interferons – Induction, Signaling, Evasion, and Application to Combat COVID-19. Cell Host & Microbe. 2020;27(6):870-878. doi:10.1016/j.chom.2020.05.008
- Platanias L.C. Mechanisms of type-I-and-type-II-interferon-mediated signalling. Nature Reviews Immunology. 2005;5(5):375-386. doi:10.1038/nri1604
- RECOVERY Collaborative Group. Baricitinib in patients admitted to hospital with COVID-19 (RECOVERY): a randomised, controlled, open-label, platform trial and updated meta-analysis. Lancet. 2022;400(10349):359-368. doi:10.1016/S0140-6736(22)01109-6
- Sabbatinelli J., et al. Decreased serum levels of the inflammaging marker miR-146a are associated with clinical non-response to tocilizumab in COVID-19 patients. Mechanisms of Ageing and Development. 2021;193:111413. doi:10.1016/j.mad.2020.111413
- Sawcer S., et al. (International Multiple Sclerosis Genetics Consortium). Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature. 2011;476(7359):214-219. doi:10.1038/nature10251
- Solé X, Guinó E, Valls J, Iniesta R, Moreno V. SNPStats: a web tool for the analysis of association studies. Bioinformatics. 2006;22(15):1928-1929. doi:10.1093/bioinformatics/btl268
- Stahl E.A., et al. Genome-wide association study meta-analysis identifies seven new rheumatoid arthritis risk loci. Nature Genetics. 2010;42(6):508-514. doi:10.1038/ng.582
- Taganov K.D., et al. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(33):12481-12486. doi:10.1073/pnas.0605298103
- Taha S.I., et al. Role of immune-inflammatory biomarkers and their derived ratio in predicting COVID-19 severity and mortality. Scientific Reports. 2025;15:39003. doi:10.1038/s41598-025-24173-7
- Zhou Z., et al. Heightened Innate Immune Responses in the Respiratory Tract of COVID-19 Patients. Cell Host & Microbe. 2020;27(6):883-890.e2. doi:10.1016/j.chom.2020.04.017
Статья поступила в редакцию 12 февраля 2026 г.
Поступила после доработки 6 марта 2026 г.
Принята к печати 20 марта 2026 г.
Received 12, February, 2026
Revised 6, March, 2026
Accepted 20, March, 2026