Введение
Изучение чешуекрылых (Lepidoptera) Ростовской области (РО) имеет более чем вековую историю, восходящую к работам конца XIX – начала XX века (Ершов, Фильд, 1870; Алфераки, 1876, 1877, 1880, 1908). Интерес к этим насекомым во многом обусловлен их хозяйственной значимостью: среди бабочек имеется много вредителей сельского и лесного хозяйства, что отражено как в ранних (Зверезомб-Зубовский, 1918, 1919), так и в последующих публикациях (Добровольский, 1939, 1950, 1959; Померанцев, 1949; Архангельский, 1929, 1931). Особое место среди них занимают листовёртки (Tortricidae) – одна из ведущих хозяйственно значимых групп чешуекрылых.
Мировая фауна Tortricidae насчитывает более 10000 видов (Zhang, 2011), в России отмечено 1170 видов (Синёв, 2019). Семейство включает множество вредителей сельского и лесного хозяйства, из которых более 100 видов обитают в Европе (Загуляев и др., 1978). Несмотря на хозяйственную значимость, фауна листовёрток Ростовской области долгое время оставалась изученной фрагментарно. В ранних работах (Алфераки, 1876, 1908; Романова, 1949, 1956) зафиксировано лишь 24 вида; последующие сборы носили спорадический характер. Исключение составила диссертация А.В. Шестаковой (1967) по яблонной плодожорке Cydia pomonella (Linnaeus, 1758) и инициативный 10-летний проект А.Н. Полтавского, завершившийся публикацией кратких сведений о 91 виде (Poltavsky, 2015).
Систематическое изучение тортрицид региона активизировалось вскоре после выхода серии работ (Полтавский, Колесник, 2015; Артохин, Полтавский, 2018, 2021; Романчук, Кладковая, 2021а, б; Романчук, Колесников, 2022). Интенсивность исследований заметно возросла (Романчук, Кладковая, 2022; Романчук и др., 2023а, б, 2024а, б). В рамках инициативы «Комплексное исследование бабочек-листовёрток Ростовской области и сопредельных территорий» опубликованы крупные эколого-фаунистические обзоры для Приазовского ботанико-географического района (ПБГР) (Слюнькова и др., 2025) и Долины Нижнего Дона (ДНД) (Шидловская и др., 2026). Завершается работа по анализу сообществ листовёрток Калитвинского БГР.
Однако до настоящего времени практически не изученным оставался Азово-Егорлыкский ботанико-географический район, представляющий собой южную часть Приазовских степей. Долина Нижнего Дона географически разделяет Северные и Южные Приазовские степи – Приазовский и Азово-Егорлыкский БГР соответственно. Если разнообразие листовёрток Приазовского района уже достаточно полно охарактеризовано (Слюнькова и др., 2025), то сведения о сообществах листовёрток Азово-Егорлыкского района оставались фрагментарными и разрозненными. Это создаёт серьёзный пробел в понимании зональных закономерностей распространения листовёрток в центральной и южной частях Ростовской области. В связи с этим цель работы – сравнение экологического и таксономического разнообразия сообществ листовёрток Азово-Егорлыкского и Приазовского ботанико-географических районов.
Место проведения исследования
Изучение отдельных ботанико-географических районов Ростовской области (по Зозулин, Пашков, 1980) позволяет глубже понять видовое разнообразие листовёрток как на исследуемой территории, так и в регионе в целом. Необходимость детального исследования Азово-Егорлыкского и Приазовского районов подтверждается данными сельскохозяйственных и аграрных исследований. Согласно оценке земель указанных БГР в рамках природно-сельскохозяйственного зонирования, бонитет почв пашни здесь является самым высоким в Ростовской области и составляет 68 и 62 балла соответственно (Косенко, 2022). Это характеризует данные территории как сельскохозяйственно значимые и обусловливает острую необходимость в тщательном изучении разнообразия и фенологии вредителей из семейства листовёрток, обитающих в этих БГР.
Азово-Егорлыкский ботанико-географический район представляет собой территорию с довольно однородным растительным покровом, сложенным преимущественно степными сообществами: разнотравно-типчаково-ковыльными, разнотравно-злаковыми и типчаковыми, с участием редких, в том числе эндемичных видов. В отличие от других степей приазовского типа, данный район отличается более ксерофильным характером флоры и обеднённостью её состава (Фишкин, Узденов, 2018). Несмотря на степной профиль, этот БГР включает в себя разнообразные стации: водно-болотные угодья, древесно-кустарниковые заросли, скверы в условиях плотной городской застройки. Подавляющее число видов и экземпляров листовёрток было собрано на участках древесно-кустарниковых насаждений и в плодово-ягодных садах.
Список пунктов сбора: г. Азов – берег реки (древесно-кустарниковые заросли); пос. Победа – агроценоз (сады в населённом пункте); пос. Каяльский – долина реки (водно-болотные угодья); село Самарское – агроценоз (сады в населённых пунктах); станица Старочеркасская – агроценоз (древесно-кустарниковые заросли); станица Кировская – агроценоз (сады в населённых пунктах); г. Зерноград – урбоценоз (сквер, древесно-кустарниковые заросли); окр. хут. Заполосный – агроценоз (сады в населённых пунктах); пос. Вороново – агроценоз (сады в населённых пунктах); село Лысогорка – долина реки (древесно-кустарниковые заросли); Веселовское водохранилище – долина реки (древесно-кустарниковые заросли); Хлебная балка – остепнённый участок (разнотравная степь); Гигант – агроценоз (древесно-кустарниковые насаждения, сады в населённом пункте), долина реки (древесно-кустарниковые заросли).
Приазовский ботанико-географический район характеризуется сочетанием степной, лугово-болотной и древесно-кустарниковой растительности. Отличается почти полным отсутствием байрачных лесов, на смену которым пришли кустарниковые формации. Встречаются балки с типичными для полосы приазовских настоящих степей сообществами – разнотравно-дерновиннозлаковыми и каменистыми разнотравно-дерновинно-злаковыми (Зозулин, Пашков, 1980; Полтавский, Артохин, 2012; Фишкин, Узденов, 2018).
Список пунктов сбора: окр. села Синявское – балка реки Донской Чулек (ковыльная степь, древесно-кустарниковые заросли); село Весело-Вознесенка – агроценоз (сады в населённом пункте); балка реки Донской Чулек – разнотравная степь, закустаренные остепнённые склоны; Кульбаково – долина реки (сухая степь); окр. хут. Морской Чулек – робиниевая лесополоса (древесно-кустарниковые заросли); остановочный пункт 1300 км – агроценоз (сады в населённом пункте); охраняемый природный объект «Каменная Балка» – остепнённый участок (остепнённые балочные склоны и овраги); УОХ ЮФУ «Недвиговка» – долина реки (древесно-кустарниковые заросли); слобода Большекрепинская – долина реки (кустарниковые склоны); г. Ростов-на-Дону, Щепкинский лес – искусственный лесной массив; г. Ростов-на-Дону, Парк им. Николая Островского – древесно-кустарниковые насаждения; г. Ростов-на-Дону, Ботанический сад ЮФУ – урбоценоз (древесно-кустарниковые заросли, степные залежи); хут. Хапры – остепнённый участок (разнотравная степь); г. Аксай – урбоценоз (сады в населённом пункте); хут. Коныгин – долина реки (низинный лес). Места сбора представленными типизированными местообитаниями (Рис. 1).

Рис. 1 – Ландшафтно-биотопический профиль некоторых исследованных участков Южных и Северных Приазовских степей (фото ориг.): а – байрачные и низинные леса; б – плодово-ягодные сады; в – древесно-кустарниковые заросли; г – древесно-кустарниковые насаждения; д –частные секторы; е – остепнённые балочные склоны и овраги; ж –закустаренные остепнённые склоны и овраги; з – разнотравные степи; и –степные залежи; к – ковыльные степи; л – водно-болотные угодья; м – скверы в плотной городской застройке
Материалы и методы
В ходе исследования были использованы все доступные литературные источники по тортрицидофауне региона. Фактический материал включал как коллекционные сборы энтомологов прошлых лет, так и оригинальные данные, собранные авторами на территории Азово-Егорлыкского и Приазовского БГР. Эти инициативные сборы значительно дополнили общий массив сведений.
Сбор материала осуществлялся несколькими способами (Рис. 2):
1) Аттрактивным с использованием клеевых ловушек (Рис. 2а) с синтетическими половыми феромонами самок Cydia nigricana, Aspila funebrana. Период экспонирования аналогичный.
2) Аттрактивным с использованием белого светоотражающего экрана, на фоне которого размещалась газоразрядная ртутно-вольфрамовая лампа прямого включения TDM Electric (Рис. 2б). Сбор с экрана привлечённых на свет листовёрток осуществлялся ручным способом в специализированные морилки. Для замаривания применялся этилацетат. Работа велась в сумеречное и ночное время. Источником электроэнергии в разных случаях служили стационарная сеть и портативная электростанция.
3) Аттрактивным с использованием ловушек-сборщиков (Рис. 2в) с диспенсерами, содержащими синтетические половые феромоны самок Aspila molesta, A. funebrana, Cydia pyrivora. Ловушки экспонировались в течение 14 дней, после чего осуществлялась их замена.
Собранный материал определён и хранится на ватных матрасиках либо в расправленном/смонтированном виде (Рис. 3б, в). При препаровке использован стандартный инструментарий. Для определения применяли методы морфологического анализа гениталий и габитусов (Рис. 3а) с привлечением интернет-баз данных и специализированной литературы (Пестициды…, https://www.pesticidy.ru; EPPO…, https://gd.eppo.int; Lepiforum…, https://lepiforum.org; Moth Dissection, https://mothdissection.co.uk; Tortricid…, http://www.tortricidae.com), а также тематическая литература (Данилевский, Кузнецов, 1968; Корнелио, 1986; Определитель…, 2001, 2016; Романчук, Колесников, 2022 и др.).

Рис. 2 – Полевые методы исследований: а – установка клеевой феромонной ловушки в Приазовском БГР (фото Р.В. Романчука); б – сбор со светоотражающего экрана в Азово-Егорлыкском БГР, пос. Гигант (фото Ю.С. Мижерицкой); в – установка феромонной ловушки в Азово-Егорлыкском БГР, хут. Бурхановка (фото Т.Е. Тихоновой); г – разбор материала после сбора со светоотражающего экрана в Приазовском БГР, УОХ ЮФУ (фото Е.А. Богаевой)
Для препаровки и изучения гениталий листовёрток (Рис. 3г) преимущественно использовали «холодный метод» – мацерацию брюшка в 10% растворе аммиака в течение нескольких дней. При необходимости срочной идентификации применяли «горячий метод» – мацерацию в кипящем растворе NaOH. Выделенные органы помещали в микропробирки с глицерином, которые подкалывали к экземплярам имаго. Часть препаратов изготовлена на предметных стёклах. Каждая единица хранения снабжена полным набором этикеток.

Рис. 3 – Камеральные методы обработки материала:
а – идентификация тортрицид по габитусу (фото Р.В. Романчука); б – модульный способ хранения насекомых (фото Т.Е. Тихоновой); в – расстановка материала в энтомологическую коробку (фото Т.Е. Тихоновой); г – изучение гениталий листовёрток с помощью бинокуляра (фото Р.В. Романчука)
Результаты определения вносились в базу данных Excel. Схема размещения сведений и характер содержания информации следующие: подсемейство; триба; род; вид; автор; год описания; область; районы, административные центры; населённые пункты, локалитеты; биотопы; стация; метод сбора; целевой организм (для феромонных ловушек); дата сбора; кол-во экз.; год первого выявления; год последнего выявления; выявленный максимум (экз.); время лёта; обилие; экологическая группа (детальный вариант классификации см. в Недошивина, 2010); трофическая приуроченность (растительное семейство); трофическая приуроченность (растительные рода); широта трофических связей; приуроченность к жизненным формам растений (по Недошивиной, 2011); основная группа вредителя по повреждаемым объектам; хозяйственное значение в текущих условиях; инвазивный вид; карантинный вид (А1/А2); источники материала.
Расчёты и графические построения выполнены в Excel. Карты распространения видов-вредителей по исследованным БГР созданы в Paint Tool SAI. Сходство сообществ листовёрток оценивали по коэффициенту Чекановского–Съёренсена в программе PAST. Видовое богатство сравнивали с помощью кривых разрежения (Сушко, 2021). Уровень разнообразия определяли по индексам Шеннона (H), Симпсона (D), Бергера–Паркера (d) и Бриллуэна (HB) (Дунаев, 1997; Шайхутдинова, 2019).
Результаты и обсуждение
Количественные данные по сборам листовёрток исследованных БГР. В период 1937–2025 гг. на Приазовском БГР отмечено 128 видов 5807 экз. Tortricidae. По Азово-Егорлыкскому БГР (2003–2025 гг.) показатели ниже: 75 видов, 914 экз. (Рис. 4).
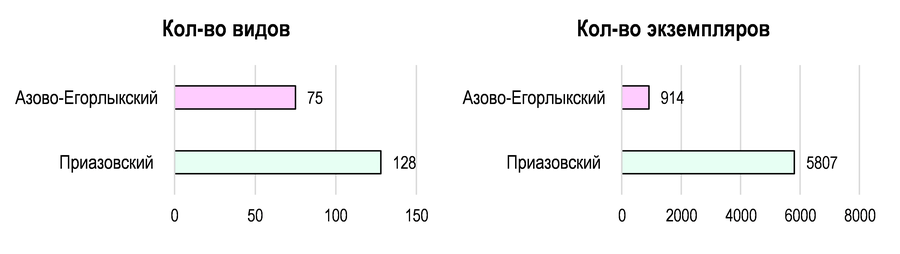
Рис. 4 – Общие количественные данные по сборам листовёрток исследованных БГР
Эти данные свидетельствуют о значительно более низкой изученности Южных Приазовских степей по сравнению с Северными. Несмотря на то, что в Приазовском БГР собрано в 8 раз больше экземпляров (5807 против 914), выявленное число видов превышает таковое для Азово-Егорлыкского БГР лишь в 2 раза (128 против 75). Формально это указывает на высокую пестроту сообществ и значительное видовое разнообразие листовёрток в Южных Приазовских степях, которое ещё предстоит выявить.
Таксономический анализ листовёрток Южных и Северных Приазовских степей. В Азово-Егорлыкском БГР за 2003–2025 гг. выявлено 75 видов листовёрток (Табл. 1). Подсемейство Tortricinae насчитывает 28 видов (37%), доминируют трибы Cochylini (16%) и Archipini (11%). Подсемейство Olethreutinae представлено 47 видами (63%), с преобладанием триб Eucosmini (28%) и Grapholitini (15%).
Таблица 1 – Таксономическая организация сообществ листовёрток Южных Приазовских степей
| Таксономическая группа | Число видов (S) | % от общего числа видов |
|---|---|---|
| Подсемейство Tortricinae | 28 | 37% |
| Триба Cochylini | 12 | 16% |
| Триба Archipini | 8 | 11% |
| Триба Cnephasiini | 4 | 5% |
| Триба Tortricini | 3 | 4% |
| Триба Ramapesiini | 1 | 1% |
| Подсемейство Olethreutinae | 47 | 63% |
| Триба Eucosmini | 21 | 28% |
| Триба Grapholitini | 11 | 15% |
| Триба Olethreutini | 7 | 9% |
| Триба Lobesiini | 3 | 4% |
| Триба Enarmoniini | 3 | 4% |
| Триба Bactrini | 2 | 3% |
| Всего | 75 |
В Приазовском БГР за 1937–2025 гг. зарегистрировано 128 видов (Табл. 2). Доля Tortricinae здесь составляет 50 видов (40%), Olethreutinae – 78 видов (60%). Структура доминирования триб сходна: в Tortricinae лидируют Cochylini (15%) и Archipini (10%), в Olethreutinae – Eucosmini (27%) и Grapholitini (14%).
Таблица 2 – Таксономическая организация сообществ листовёрток Северных Приазовских степей
| Таксономическая группа | Число видов (S) | % от общего числа видов |
|---|---|---|
| Подсемейство Tortricinae | 50 | 40% |
| Триба Cochylini | 17 | 15% |
| Триба Archipini | 14 | 10% |
| Триба Cnephasiini | 10 | 8% |
| Триба Tortricini | 7 | 5% |
| Триба Ramapesiini | 2 | 2% |
| Подсемейство Olethreutinae | 78 | 60% |
| Триба Eucosmini | 36 | 27% |
| Триба Grapholitini | 19 | 14% |
| Триба Olethreutini | 9 | 7% |
| Триба Enarmoniini | 5 | 5% |
| Триба Endotheniini | 5 | 4% |
| Триба Lobesiini | 3 | 2% |
| Триба Bactrini | 1 | 1% |
| Всего | 128 |
Значительное различие между БГР по числу видов и объёму сборов объясняется, прежде всего, неодинаковой интенсивностью исследований. Активная энтомологическая работа на территории УОХ ЮФУ «Недвиговка», в окрестных ООПТ и вдоль тракта Ростов‑на‑Дону–Таганрог – исторически сложившаяся данность, закономерно приведшая к большей репрезентативности материала по Северным Приазовским степям.
Анализ относительного обилия листовёрток Южных и Северных Приазовских степей. Распределение тортрицид по рассматриваемым ниже условным группам и общий анализ относительного обилия выполнены в соответствии с предыдущими работами (Романчук и др., 2024; Слюнькова и др., 2025). В южных Приазовских степях группа «очень редкие» включает 39 видов (52%) (Рис. 5): Phtheochroa farinosana, Ph. fulvicinctana, Phalonidia albipalpana, Ph. manniana, Aethes williana, Ae. nefandana, Ae. tesserana, Brevicornutia pallidana, Acleris aspersana, Cnephasia genitalana, Argyrotaenia ljungiana, Clepsis neglectana, Lozotaeniodes formosana, Bactra lancealana, B. lacteana, Celypha doubledayana, Lobesia botrana, L. artemisiana, Thiodia citrana, Epinotia tenerana, Pelochrista fusculana, P. metria, P. caecimaculana, P. subtiliana, P. labyrinthicana, Eucosma luciana, Epibactra immundana, Gypsonoma minutana, Epiblema sticticana, E. cirsiana, Rhyacionia pinivorana, Enarmonia formosana, Ancylis unculana, A. mitterbacheriana, Cydia splendana, Pammene suspectana, Dichrorampha plumbana, D. vancouverana, D. agilana.
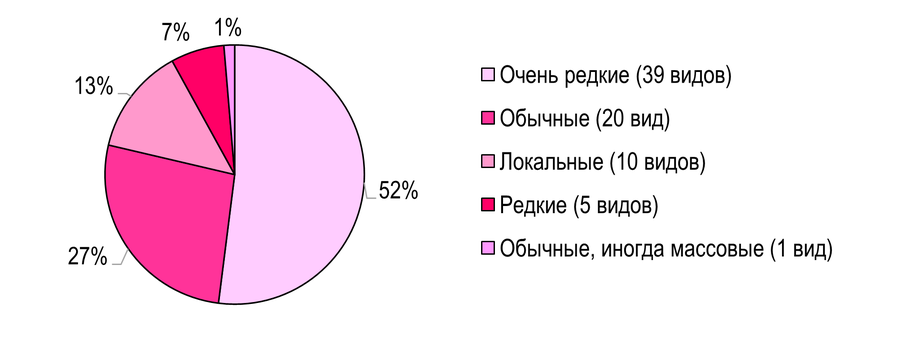
Рис. 5 – Анализ относительного обилия листовёрток Азово-Егорлыкского БГР
К группе «обычные» отнесено 20 видов: Phalonidia contractana, Agapeta hamana, Tortrix viridana, Aleimma loeflingiana, Cnephasia stephensiana, C. communana, Archips podana, A. xylosteana, A. rosana, Clepsis spectrana, C. pallidana, Hedya nubiferana, Celypha striana, C. rosaceana, Olethreutes arcuella, Epiblema scutulana, Aspila funebrana, Cydia pyrivora, C. fagiglandana, C. amplana.
Группа «локальные» включает 10 видов: Eugnosta magnificana, Pontoturania posterana, Cnephasia incertana, Periclepsis cinctana, Hedya pruniana, Celypha cespitana, Pelochrista arabescana, Eucosma cana, E. conterminana, Aspila molesta.
«Редкие» – 5 видов: Lobesia abscisana, Eucosma pupillana, Epiblema costipunctana, E. graphana, E. sarmatana. Cydia pomonella – обычный, иногда массовый вид.
Для Северных Приазовских степей группа «очень редкие» представлена 68 видами (Рис. 6): Phtheochroa pulvillana, Ph. fulvicinctana, Cochylimorpha woliniana, C. obliquana, C. subwoliniana, Diceratura roseofasciana, Phalonidia albipalpana, Aethes beatricella, Ae. bilbaensis, Cochylidia implicitana, Neocochylis hybridella, Brevicornutia pallidana, Acleris rhombana, A. kochiella, Neosphaleroptera nubilana, Cnephasia alticolana, C. asseclana, C. genitalana, Epagoge grotiana, Paramesia gnomana, Choristoneura diversana, Ch. lafauryana, Argyrotaenia ljungiana, Pandemis dumetana, Clepsis neglectana, Bactra lancealana, Endothenia pullana, E. nigricostana, Hedya ochroleucana, Lobesiodes euphorbiana, L. botrana, Thiodia citrana, Th. irinae, Epinotia festivana, Zeiraphera ratzeburgiana, Z. isertana, Pelochrista caecimaculana, P. mollitana, P. subtiliana, P. infidana, Eucosma flavispecula, E. aemulana, E. tundrana, E. wimmerana, Gypsonoma minutana, G. sociana, Epiblema sticticana, E. cirsiana, Notocelia cynosbatella, N. tetragonana, N. roborana, N. trimaculana, Rhyacionia buoliana, Ancylis comptana, A. unculana, A. apicella, A. mitterbacheriana, Aspila janthinana, Grapholita compositella, G. delineana, G. caecana, Ephippiphora gemmiferana, Cydia servillana, C. splendana, Pammene suspectana, Dichrorampha simpliciana, D. incognitana, D. agilana.

Рис. 6 – Анализ относительного обилия листовёрток Приазовского БГР
К группе «обычные» отнесено 28 видов: Phalonidia contractana, Agapeta hamana, Tortrix viridana, Aleimma loeflingiana, Cnephasia stephensiana, C. pasiuana, C. communana, Archips podana, A. crataegana, A. xylosteana, A. rosana, Aphelia ferugana, Clepsis spectrana, C. pallidana, Endothenia quadrimaculana, Hedya salicella, H. nubiferana, Celypha striana, C. rosaceana, Olethreutes arcuella, Epiblema scutulana, E. foenella, Notocelia uddmanniana, Aspila funebrana, Cydia nigricana, C. pyrivora, C. fagiglandana, C. amplana.
Группа «редкие» включает 17 видов: Eugnosta lathoniana, Acleris holmiana, A. forsskaleana, Endothenia gentianaeana, E. marginana, Celypha flavipalpana, Lobesia abscisana, Thiodia torridana, Spilonota ocellana, S. laricana, Eucosma albidulana, E. pupillana, Epiblema costipunctana, E. graphana, E. sarmatana, Ancylis achatana, Grapholita coronillana.
«Локальные» представлены 14 видами: Eugnosta magnificana, Pontoturania posterana, Acleris variegana, Doloploca punctulana, Eana incanana, Cnephasia incertana, Choristoneura hebenstreitella, Ptycholoma lecheana, Hedya pruniana, Celypha cespitana, Eucosma cana, E. metzneriana, E. conterminana, Aspila molesta. Cydia pomonella – обычный, иногда массовый вид.
В сборах из обоих БГР доминируют виды, отнесённые к группе «очень редкие». Это обусловлено интенсификацией изучения листовёрток в последние годы, что привело к значительному выявлению новых для Ростовской области видов. Поскольку первоначально такие находки, как правило, ограничиваются единичными экземплярами, видам по умолчанию присваивается статус «очень редкий». В дальнейшем, при повторных обнаружениях и накоплении данных о распространении в регионе, эти виды могут быть переведены в другие категории.
Соотношение «обычных» и «локальных» видов в обоих БГР сходно. При этом на территории Северных Приазовских степей выявлено в три раза больше редких видов, чем в Южных.
Распределение листовёрток по экологическим группам. Доминантной экологической группой для Азово-Егорлыкского БГР являются лесные мезофилы – 24 вида (32%) (Рис. 7): Tortrix viridana, Aleimma loeflingiana, Cnephasia stephensiana, Archips podana, A. rosana, Clepsis spectrana, Lozotaeniodes formosana, Hedya nubiferana, H. pruniana, Epinotia tenerana, Eucosma cana, Gypsonoma minutana, Rhyacionia pinivorana, Enarmonia formosana, Ancylis unculana, A. mitterbacheriana, Aspila funebrana, A. molesta, Cydia pomonella, C. pyrivora, C. splendana, C. fagiglandana, C. amplana, Pammene suspectana.
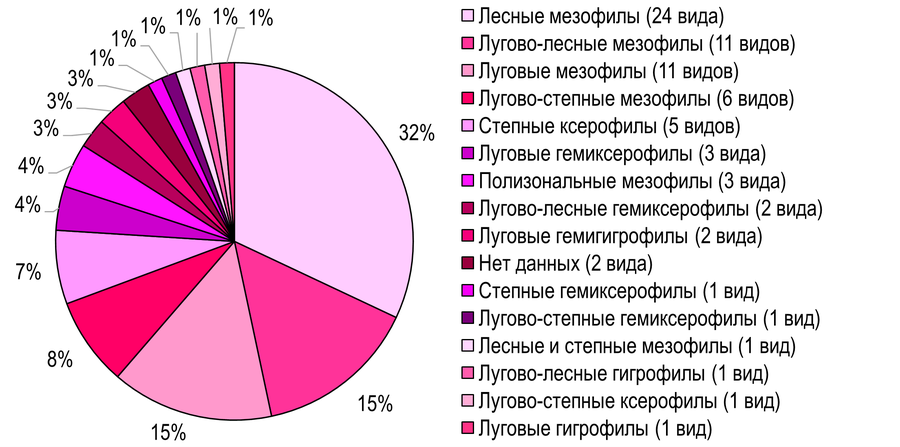
Рис. 7 – Распределение листовёрток по экологическим группам на территории Южных Приазовских степей
Субдоминантами выступают лугово-лесные мезофилы (11 видов, 15%): Phalonidia contractana, Agapeta hamana, Aethes williana, Pontoturania posterana, Cnephasia genitalana, Archips xylosteana, Clepsis pallidana, Celypha striana, C. rosaceana, Olethreutes arcuella, Lobesia botrana. А также луговые мезофилы (11 видов, 15%) (Рис. 11): Phtheochroa fulvicinctana, Phalonidia albipalpana, Aethes tesserana, Acleris aspersana, Cnephasia incertana, Clepsis neglectana, Bactra lancealana, Celypha cespitana, Pelochrista caecimaculana, Eucosma conterminana, E. luciana.
Доминантная экологическая группа в Приазовском БГР – лесные мезофилы (49 видов; 38%) (Рис. 8): Tortrix viridana, Aleimma loeflingiana, Acleris holmiana, A. forsskaleana, A. rhombana, A. variegana, A. kochiella, Neosphaleroptera nubilana, Eana incanana, Cnephasia stephensiana, Archips podana, A. crataegana, A. rosana, Choristoneura diversana, Ch. hebenstreitella, Ptycholoma lecheana, Pandemis dumetana, Clepsis spectrana, Endothenia marginana, E. quadrimaculana, Hedya salicella, H. nubiferana, H. pruniana, H. ochroleucana, Spilonota ocellana, S. laricana, Epinotia festivana, Zeiraphera isertana, Eucosma cana, Gypsonoma minutana, G. sociana, Notocelia cynosbatella, N. tetragonana, N. uddmanniana, Rhyacionia buoliana, Ancylis unculana, A. apicella, A. achatana, A. mitterbacheriana, Aspila funebrana, A. janthinana, A. molesta, Cydia pomonella, C. pyrivora, C. servillana, C. splendana, C. fagiglandana, C. amplana, Pammene suspectana.
Субдоминанта – лугово-лесные мезофилы (22 вида; 17%): Cochylimorpha subwoliniana, Phalonidia contractana, Agapeta hamana, Neocochylis hybridella, Pontoturania posterana, Doloploca punctulana, Cnephasia genitalana, Epagoge grotiana, Archips xylosteana, Aphelia ferugana, Clepsis pallidana, Endothenia nigricostana, Celypha striana, C. rosaceana, C. flavipalpana, Olethreutes arcuella, Lobesia botrana, Zeiraphera ratzeburgiana, Notocelia roborana, N. trimaculana, Grapholita delineana, G. coronillana.

Рис. 8 – Распределение листовёрток по экологическим группам на территории Северных Приазовских степей
Третья по численности группа – луговые мезофилы (19 видов; 15%): Phtheochroa fulvicinctana, Diceratura roseofasciana, Phalonidia albipalpana, Cnephasia incertana, C. alticolana, Clepsis neglectana, Bactra lancealana, Endothenia pullana, Celypha cespitana, Lobesiodes euphorbiana, Pelochrista caecimaculana, Eucosma flavispecula, E. conterminana, Grapholita compositella, G. caecana, Ephippiphora gemmiferana, Cydia nigricana, Dichrorampha simpliciana, D. incognitana.
Распределение листовёрток по экологическим группам на территории Азово-Егорлыкского и Приазовского БРГ демонстрируют сходную картину: с заметным перевесом в сообществах доминируют лесные мезофильные виды (Рис. 7, 8), что весьма характерно для тортрицид с учётом экологических преферендумов большинства из них (Романчук, Кладковая, 2021а, б; Романчук и др., 2024, 2025; Слюнькова и др., 2025). В обоих БГР субдоминантами выступают лугово-лесные мезофилы, третью позицию занимают луговые мезофилы. Полученные данные свидетельствуют о наличии в региональной фауне тортрицид лесо-лугового мезофильного экологического субкомплекса, сформированного большинством видов.
Трофическая приуроченность и хозяйственное значение листовёрток Южных и Северных Приазовских степей.
Трофическая приуроченность. Результаты анализа трофических преферендумов листовёрток Азово-Егорлыкского и Приазовского БГР свидетельствуют о широкой кормовой базе: значительное число видов, являясь полифагами, питается на представителях нескольких семейств растений. Основу составляют Asteraceae и Rosaceae (Рис. 9, 10), среди которых присутствует много культурных видов.
В Азово-Егорлыкском БГР с Asteraceae трофически ассоциировано 30 видов тортрицид (Рис. 9): Phtheochroa farinosana, Phalonidia contractana, Agapeta hamana, Aethes tesserana, Pontoturania posterana, Cnephasia stephensiana, C. genitalana, C. communana, Argyrotaenia ljungiana, Clepsis spectrana, C. pallidana, Celypha rosaceana, Lobesia abscisana, L. artemisiana, Thiodia citrana, Pelochrista caecimaculana, P. subtiliana, P. arabescana, P. labyrinthicana, Eucosma cana, E. conterminana, E. pupillana, Epiblema sticticana, E. scutulana, E. cirsiana, E. costipunctana, E. graphana, Dichrorampha plumbana, D. vancouverana, D. agilana. С Rosaceae – 17 видов: Acleris aspersana, Archips podana, A. xylosteana, A. rosana, Argyrotaenia ljungiana, Clepsis spectrana, C. pallidana, C. neglectana, Hedya nubiferana, H. pruniana, Celypha cespitana, Lobesia botrana, Enarmonia formosana, Aspila funebrana, A. molesta, Cydia pomonella, C. pyrivora.
В Приазовском БГР с Asteraceae трофически связано 52 вида (Рис. 10): Cochylimorpha woliniana, C. obliquana, C. subwoliniana, Phalonidia contractana, Agapeta hamana, Eugnosta lathoniana, Cochylidia implicitana, Neocochylis hybridella, Pontoturania posterana, Eana incanana, Cnephasia stephensiana, C. alticolana, C. asseclana, C. pasiuana, C. genitalana, C. communana, Epagoge grotiana, Paramesia gnomana, Choristoneura diversana, Ch. lafauryana, Argyrotaenia ljungiana, Pandemis dumetana, Clepsis spectrana, C. pallidana, Celypha rosaceana, Lobesia abscisana, Thiodia citrana, Th. irinae, Th. torridana, Pelochrista caecimaculana, P. mollitana, P. subtiliana, P. infidana, Eucosma cana, E. flavispecula, E. aemulana, E. albidulana, E. metzneriana, E. tundrana, E. wimmerana, E. conterminana, E. pupillana, Epiblema sticticana, E. scutulana, E. cirsiana, E. foenella, E. costipunctana, E. graphana, Dichrorampha simpliciana, D. incognitana, D. agilana. С Rosaceae – 37 видов: Acleris holmiana, A. rhombana, A. variegana, Neosphaleroptera nubilana, Epagoge grotiana, Archips podana, A. crataegana, A. xylosteana, A. rosana, Choristoneura diversana, Ch. hebenstreitella, Ch. lafauryana, Argyrotaenia ljungiana, Ptycholoma lecheana, Pandemis dumetana, Clepsis spectrana, C. pallidana, C. neglectana, Hedya nubiferana, H. pruniana, H. ochroleucana, Celypha cespitana, Lobesia botrana, Spilonota ocellana, Zeiraphera isertana, Notocelia cynosbatella, N. tetragonana, N. uddmanniana, N. roborana, N. trimaculana, Ancylis comptana, A. achatana, Aspila funebrana, A. janthinana, A. molesta, Cydia pomonella, C. pyrivora.
На территории обоих БГР в трофическом спектре в качестве субдоминант выступают семейства Fabaceae, Fagaceae, Betulaceae и Ericaceae. При этом в Южных Приазовских степях четвёртое место по числу связанных с ними видов листовёрток занимает Betulaceae, тогда как в Северных – Fagaceae.
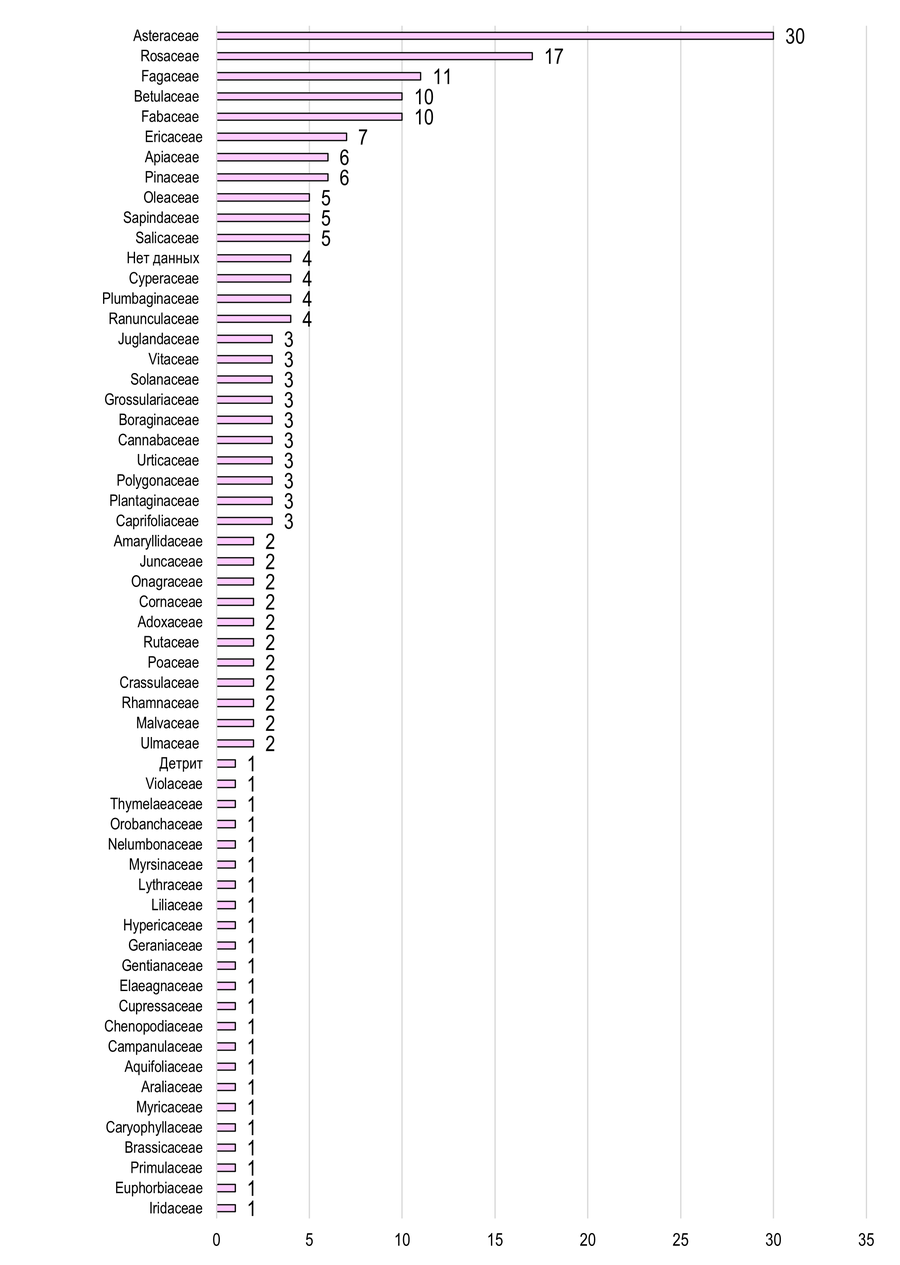
Рис. 9 – Кормовая база листовёрток Азово-Егорлыкского БГР
(с указанием числа трофически ассоциированных видов бабочек)
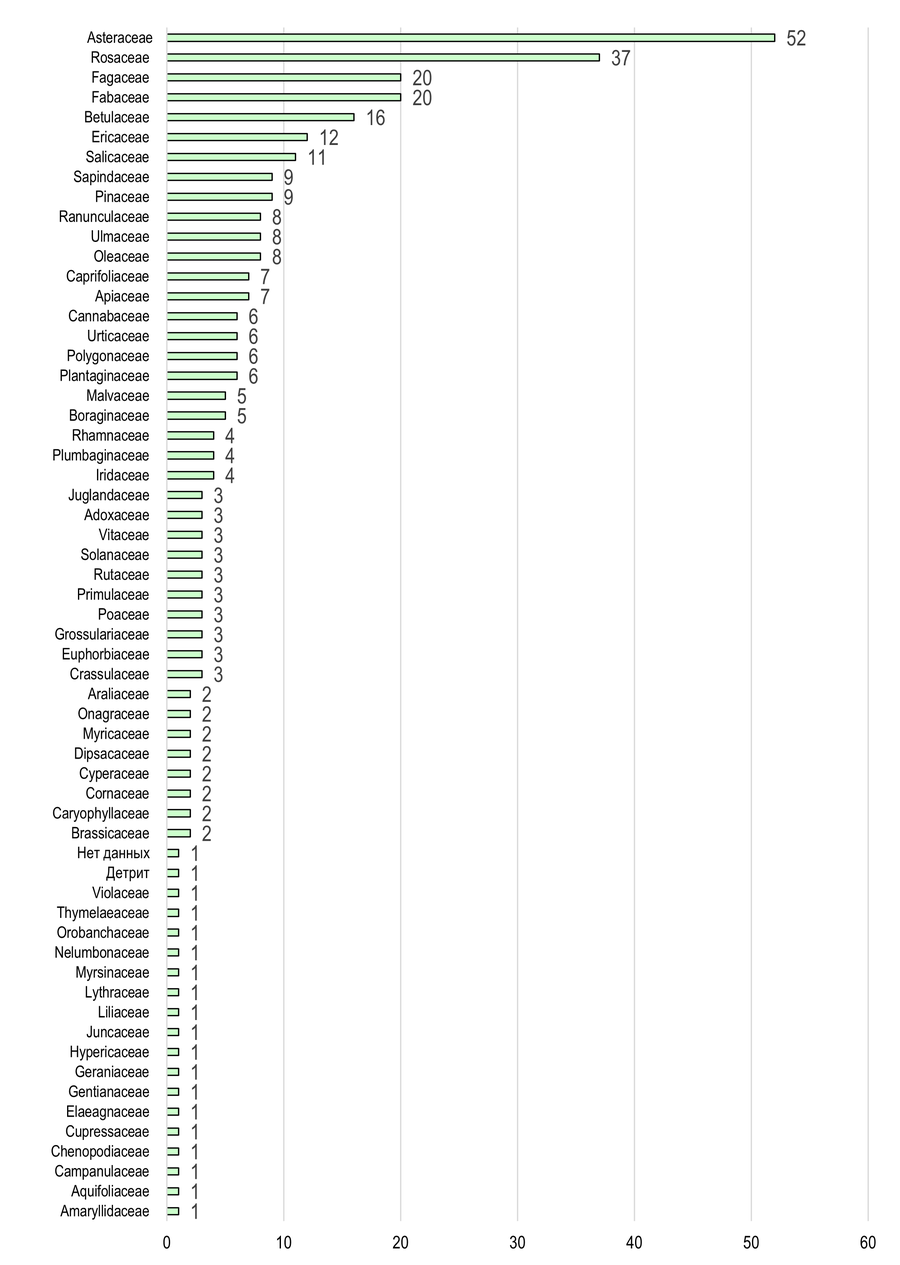
Рис. 10 – Кормовая база листовёрток Приазовского БГР
(с указанием числа трофически ассоциированных видов бабочек)
Широта трофических связей. В Азово-Егорлыкском БГР наиболее многочисленной трофической группой являются полифаги – 27 видов (36%) (Рис. 11): Phalonidia manniana, Tortrix viridana, Aleimma loeflingiana, Acleris aspersana, Cnephasia incertana, C. stephensiana, C. genitalana, C. communana, Archips podana, A. xylosteana, A. rosana, Argyrotaenia ljungiana, Clepsis spectrana, C. pallidana, Bactra lancealana, B. lacteana, Hedya pruniana, Celypha striana, C. cespitana, C. doubledayana, Olethreutes arcuella, Lobesia botrana, L. abscisana, L. artemisiana, Cydia pomonella, C. splendana, C. amplana.

Рис. 11 – Широта трофических связей листовёрток Южных Приазовских степей
Субдоминантой выступают широкие олигофаги – 24 вида (32%): Phalonidia contractana, Agapeta hamana, Aethes williana, Ae. tesserana, Pontoturania posterana, Periclepsis cinctana, Hedya nubiferana, Celypha rosaceana, Thiodia citrana, Epinotia tenerana, Eucosma cana, Epiblema sticticana, E. scutulana, E. cirsiana, E. graphana, Rhyacionia pinivorana, Enarmonia formosana, Ancylis unculana, A. mitterbacheriana, Aspila molesta, Cydia fagiglandana, Dichrorampha plumbana, D. vancouverana, D. agilana.
Менее распространены монофаги – 12 видов (16%): Phtheochroa farinosana, Ph. fulvicinctana, Phalonidia albipalpana, Brevicornutia pallidana, Clepsis neglectana, Lozotaeniodes formosana, Pelochrista caecimaculana, P. subtiliana, P. labyrinthicana, Eucosma pupillana, Epiblema costipunctana, Pammene suspectana.
Аналогично Азово-Егорлыкскому БГР, доминирующей трофической группой в Приазовском БГР являются полифаги – 46 видов (36%) (Рис. 12): Cochylimorpha subwoliniana, Tortrix viridana, Aleimma loeflingiana, Acleris variegana, Doloploca punctulana, Eana incanana, Cnephasia incertana, C. stephensiana, C. alticolana, C. asseclana, C. pasiuana, C. genitalana, C. communana, Epagoge grotiana, Paramesia gnomana, Archips podana, A. crataegana, A. xylosteana, A. rosana, Choristoneura diversana, Ch. hebenstreitella, Ch. lafauryana, Argyrotaenia ljungiana, Ptycholoma lecheana, Pandemis dumetana, Aphelia ferugana, Clepsis spectrana, C. pallidana, Bactra lancealana, Endothenia gentianaeana, E. marginana, E. quadrimaculana, Hedya pruniana, Celypha striana, C. flavipalpana, C. cespitana, Olethreutes arcuella, Lobesia botrana, L. abscisana, Spilonota ocellana, Zeiraphera isertana, Notocelia cynosbatella, Ancylis comptana, Cydia pomonella, C. splendana, C. amplana.
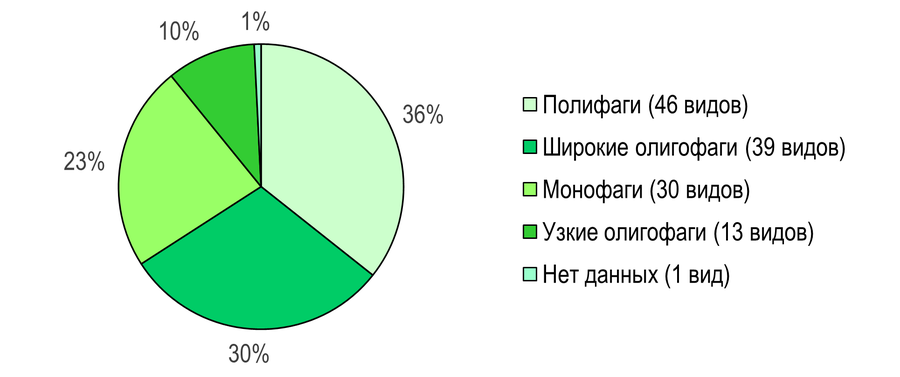
Рис. 12 – Широта трофических связей листовёрток Северных Приазовских степей
Широкие олигофаги представлены 39 видами (30%): Phalonidia contractana, Agapeta hamana, Aethes beatricella, Ae. bilbaensis, Cochylidia implicitana, Neocochylis hybridella, Pontoturania posterana, Acleris holmiana, A. rhombana, Neosphaleroptera nubilana, Endothenia nigricostana, Hedya salicella, H. nubiferana, H. ochroleucana, Celypha rosaceana, Thiodia citrana, Zeiraphera ratzeburgiana, Eucosma cana, E. aemulana, E. albidulana, Gypsonoma sociana, Epiblema sticticana, E. scutulana, E. cirsiana, E. graphana, Notocelia roborana, N. trimaculana, Ancylis unculana, A. apicella, A. achatana, A. mitterbacheriana, Aspila janthinana, A. molesta, Grapholita compositella, G. delineana, G. caecana, Cydia nigricana, C. fagiglandana, Dichrorampha agilana.
Монофаги – 30 видами (23%): Phtheochroa pulvillana, Phtheochroa fulvicinctana, Cochylimorpha woliniana, C. obliquana, Diceratura roseofasciana, Phalonidia albipalpana, Eugnosta lathoniana, Brevicornutia pallidana, Acleris kochiella, Clepsis neglectana, Endothenia pullana, Lobesiodes euphorbiana, Thiodia irinae, Th. torridana, Pelochrista caecimaculana, P. mollitana, P. subtiliana, P. infidana, Eucosma flavispecula, E. tundrana, E. wimmerana, E. pupillana, Epiblema costipunctana, Notocelia tetragonana, N. uddmanniana, Grapholita coronillana, Cydia servillana, Pammene suspectana, Dichrorampha simpliciana, D. incognitana.
Распределение листовёрток по трофическим группам сходно в обоих исследованных БГР. При этом наличие в кормовом спектре одного или двух видов растений может указывать не столько на монофагию как таковую, сколько на недостаточную изученность биологии конкретных видов.
Приуроченность к жизненным формам растений. В Азово-Егорлыкском БГР ведущей группой являются хортофаги – 38 видов (51%) (Рис. 13): Phtheochroa farinosana, Ph. fulvicinctana, Phalonidia albipalpana, Ph. manniana, Ph. contractana, Agapeta hamana, Aethes williana, Ae. nefandana, Ae. tesserana, Brevicornutia pallidana, Pontoturania posterana, Cnephasia incertana, C. stephensiana, C. genitalana, C. communana, Clepsis neglectana, Bactra lancealana, B. lacteana, Celypha rosaceana, Lobesia abscisana, L. artemisiana, Thiodia citrana, Pelochrista caecimaculana, P. subtiliana, P. arabescana, P. labyrinthicana, Eucosma cana, E. conterminana, E. pupillana, E. luciana, Epiblema sticticana, E. scutulana, E. cirsiana, E. costipunctana, E. graphana, Dichrorampha plumbana, D. vancouverana, D. agilana.

Рис. 13 – Приуроченность листовёрток к жизненным формам растений в Южных Приазовских степях
В соподчинённом положении находятся дендрофаги – 17 видов (23%): Aleimma loeflingiana, Archips xylosteana, A. rosana, Lozotaeniodes formosana, Olethreutes arcuella, Gypsonoma minutana, Rhyacionia pinivorana, Ancylis mitterbacheriana, Aspila funebrana, A. molesta, Cydia pomonella, C. pyrivora, C. splendana, C. fagiglandana, C. amplana, Pammene suspectana.
В Приазовском БГР доминирующей группой также являются хортофаги, составляющие половину известного видового состава листовёрток данной территории – 64 вида (50%) (Рис. 14): Phtheochroa pulvillana, Ph. fulvicinctana, Cochylimorpha woliniana, C. subwoliniana, Diceratura roseofasciana, Phalonidia albipalpana, Ph. contractana, Agapeta hamana, Eugnosta lathoniana, Aethes beatricella, Ae. bilbaensis, Cochylidia implicitana, Neocochylis hybridella, Brevicornutia pallidana, Pontoturania posterana, Eana incanana, Cnephasia incertana, C. stephensiana, C. alticolana, C. asseclana, C. pasiuana, C. genitalana, C. communana, Clepsis neglectana, Bactra lancealana, Endothenia gentianaeana, E. marginana, E. pullana, E. nigricostana, E. quadrimaculana, Celypha rosaceana, Lobesiodes euphorbiana, Lobesia abscisana, Thiodia citrana, Th. irinae, Th. torridana, Pelochrista caecimaculana, P. mollitana, P. subtiliana, P. infidana, Eucosma cana, E. flavispecula, E. aemulana, E. albidulana, E. metzneriana, E. tundrana, E. wimmerana, E. conterminana, E. pupillana, Epiblema sticticana, E. scutulana, E. cirsiana, E. foenella, E. costipunctana, E. graphana, Grapholita compositella, G. delineana, G. coronillana, G. caecana, Ephippiphora gemmiferana, Cydia nigricana, Dichrorampha simpliciana, D. incognitana, D. agilana.

Рис. 14 – Приуроченность листовёрток к жизненным формам растений в Северных Приазовских степях
Субдоминантой выступают дендрофаги – 28 видов (22%): Aleimma loeflingiana, Acleris forsskaleana, A. kochiella, Archips crataegana, A. xylosteana, A. rosana, Ptycholoma lecheana, Hedya salicella, Olethreutes arcuella, Spilonota ocellana, S. laricana, Epinotia festivana, Zeiraphera ratzeburgiana, Z. isertana, Gypsonoma minutana, G. sociana, Rhyacionia buoliana, Ancylis mitterbacheriana, Aspila funebrana, A. janthinana, A. molesta, Cydia pomonella, C. pyrivora, C. servillana, C. splendana, C. fagiglandana, C. amplana, Pammene suspectana.
Азово-Егорлыкский и Приазовский ботанико-географические районы имеют общее ядро видов, приуроченных к определённым жизненным формам растений. Это ядро включает хортофагов, дендрофагов и дендро-тамнофагов. В соподчинённом положении находятся такие группы, как тамнофаги, тамно-хортофаги и хорто-тамнофаги. Для Северных Приазовских степей характерно наличие большего разнообразия «переходных» групп ввиду присутствия на данной территории более разнообразных растительных сообществ.
Распределение по основным группам вредителей. В Южных Приазовских степях основу сообществ листовёрток составляют виды, не имеющие хозяйственного значения (47 видов; 63%). Среди вредителей доминируют вредители леса – 10 видов (14%) (Рис. 15): Tortrix viridana, Aleimma loeflingiana, Lozotaeniodes formosana, Epinotia tenerana, Gypsonoma minutana, Rhyacionia pinivorana, Ancylis unculana, Cydia splendana, C. fagiglandana, C. amplana.
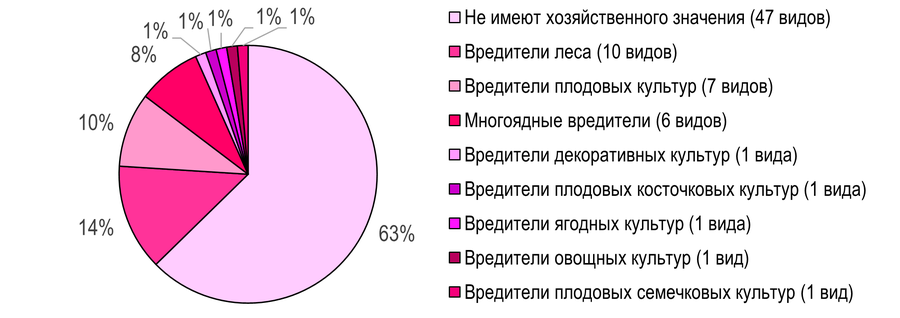
Рис. 15 – Распределение листовёрток по основным биологическим группам вредителей на территории Азово-Егорлыкского БГР
В меньшей степени представлены вредители плодовых культур (7 видов; 14%): Acleris aspersana, Archips podana, A. xylosteana, Hedya nubiferana, Enarmonia formosana, Aspila molesta, Cydia pomonella.
В Северных Приазовских степях также преобладают виды без хозяйственного значения (59%). Вредители леса насчитывают 16 видов (12%) (Рис. 16): Tortrix viridana, Aleimma loeflingiana, Acleris forsskaleana, A. kochiella, Hedya salicella, Spilonota ocellana, Epinotia festivana, Zeiraphera ratzeburgiana, Gypsonoma minutana, G. sociana, Rhyacionia buoliana, Ancylis unculana, A. apicella, Cydia splendana, C. fagiglandana, C. amplana.
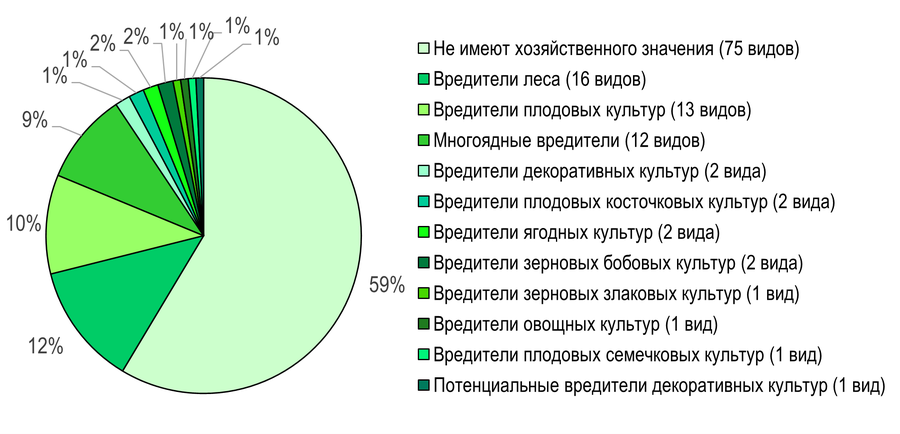
Рис. 16 – Распределение листовёрток по основным биологическим группам вредителей на территории Приазовского БГР
Вредители плодовых культур представлены 13 видами (10%): Acleris holmiana, A. rhombana, Neosphaleroptera nubilana, Archips podana, A. crataegana, A. xylosteana, Choristoneura diversana, Hedya nubiferana, H. ochroleucana, Notocelia trimaculana, Ancylis achatana, Aspila molesta, Cydia pomonella.
В обоих БГР доминируют не имеющие хозяйственного значения виды, многие из которых являются хортофагами, питающимися на дикорастущих травах. Ядро хозяйственно значимых видов образуют вредители леса, плодовых культур и многоядные формы. Различия в их численности между Южными и Северными Приазовскими степями, вероятно, обусловлены неодинаковой интенсивностью и полнотой исследований.
Основные виды листовёрток вредителей исследованных БГР.
Cnephasia stephensiana (Doubleday, 1849) – обычный лесной мезофил. Полифаг, хортофаг (Asteraceae, Fabaceae, Plantaginaceae), вредитель декоративных культур. Время лёта: май, июнь, июль, август. Точки сбора в Азово-Егорлыкском БГР: Гигант (агроценоз, сады в населённом пункте), Зерноград (урбоценоз, древесно-кустарниковые заросли), Победа (агроценоз, сады в населённом пункте) (Рис. 17а, 18а).
Hedya nubiferana (Haworth, 1811) – обычный лесной мезофил. Широкий олигофаг, дендро-тамнофаг (Rosaceae: Malus, Nok, Prunus, Crataegus, Cotoneaster, Sorbus), вредитель плодовых культур. Время лёта: май – август. Точки сбора в Азово-Егорлыкском БГР: Лысогорка (долина реки, древесно-кустарниковые заросли), Гигант (агроценоз, сады в населённом пункте), Победа (агроценоз, сады в населённом пункте) (Рис. 17б, 18б). Точки сбора в Приазовском БГР: Большекрепинская (кустарниковые склоны), Коныгин (низинный лес), Ростов-на-Дону, Ботанический сад ЮФУ (древесно-кустарниковые заросли), Ростов-на-Дону, Щепкинский лес (древесно-кустарниковые заросли), УОХ ЮФУ «Недвиговка» (древесно-кустарниковые заросли), Синявское (балка реки Донской Чулек) (разнотравная степь) (Рис. 17б, 18в).
Aspila funebrana (Treitschke, 1835) – обычный лесной мезофил. Узкий олигофаг, дендрофаг (Rosaceae: Prunus spp.), вредитель плодовых косточковых культур. Время лёта: апрель, май, июнь, июль, август, сентябрь. Точки сбора в Азово-Егорлыкском БГР: Гигант (агроценоз, сады в населённом пункте), Победа (агроценоз, сады в населённом пункте), Азов (берег реки, древесно-кустарниковые заросли). Точки сбора в Приазовском БГР: Ростов-на-Дону, Ботанический сад ЮФУ (древесно-кустарниковые заросли) Советка (сад в населённом пункте), Ростов-на-Дону, Парк им. Николая Островского (древесно-кустарниковые насаждения), Синявское (балка реки Донской Чулек) (древесно-кустарниковые заросли), Морской Чулек (древесно-кустарниковые заросли), УОХ ЮФУ «Недвиговка» (древесно-кустарниковые заросли) (Рис. 17в, 18 г, д).
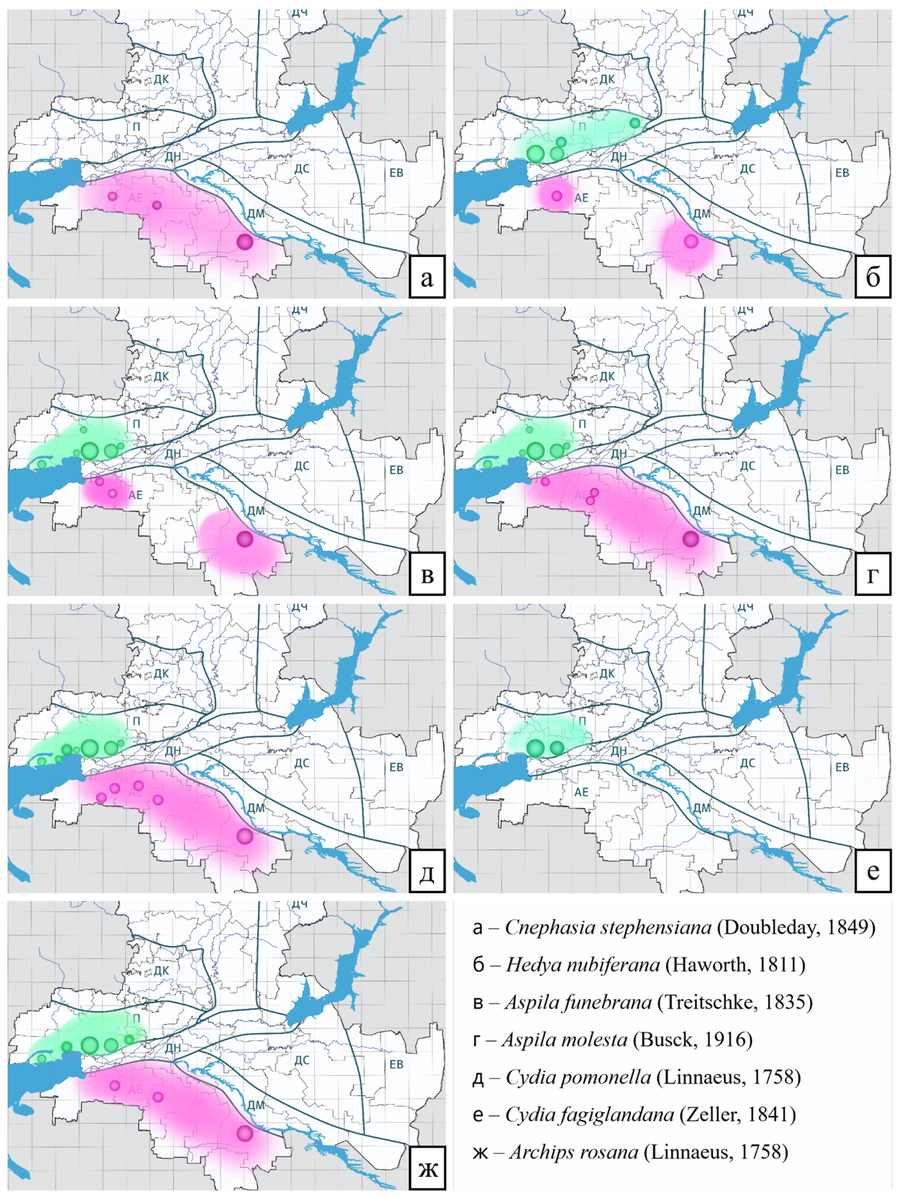
Рис. 17 – Предполагаемая зона вредоносности основных видов вредителей АЕ БГР (розовым цветом) и П БГР (зеленым цветом)
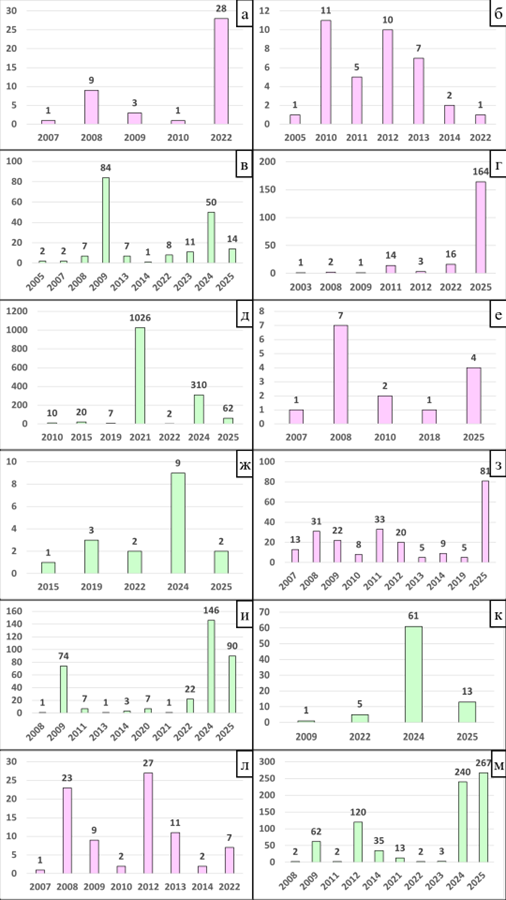
Рис. 18 – Динамика выявления основных видов вредителей АЕ БГР (розовым цветом) и П БГР (зеленым цветом): a – Cnephasia stephensiana; б, в – Hedya nubiferana; г, д – Aspila funebrana; е, ж – Aspila molesta; з, и – Cydia pomonella; к – Cydia fagiglandana; л, м –Archips rosana
Aspila molesta (Busck, 1916) – локальный лесной мезофил. Широкий олигофаг, дендрофаг (Rosaceae: Prunus, Malus, Pyrus, Cydonia, Chaenomeles, Photinia), карантинный вредный организм, вредитель плодовых культур. Время лёта: март – сентябрь. Точки сбора в Азово-Егорлыкском БГР: Гигант (агроценоз, сады в населённом пункте), Каяльский (долина реки, водно-болотные угодья), Азов (берег реки, древесно-кустарниковые заросли), Кировская (агроценоз, сады в населённом пункте). Точки сбора в Приазовском БГР: Ростов-на-Дону, Ботанический сад ЮФУ (древесно-кустарниковые заросли), Советка (сад в населённом пункте), Ростов-на-Дону, Парк им. Николая Островского (древесно-кустарниковые насаждения), Синявское (балка реки Донской Чулек) (древесно-кустарниковые заросли) (Рис. 17г, 18е, ж).
Cydia pomonella (Linnaeus, 1758) – обычный лесной мезофил. Полифаг, дендрофаг (Rosaceae, Betulaceae, Fagaceae), вредитель плодовых культур. Время лёта: май – сентябрь. Точки сбора в Азово-Егорлыкском БГР: Гигант (агроценоз, сады в населённом пункте), Зерноград (урбоценоз, сквер), Самарское (агроценоз, сады в населённых пунктах), Кировская (агроценоз, сады), Бурхановка (агроценоз, сады в населённых пунктах). Точки сбора в Приазовском БГР: Ростов-на-Дону, Ботанический сад ЮФУ (древесно-кустарниковые заросли), Таганрог (частный сектор), Русский Колодец (фруктовый сад), УОХ ЮФУ «Недвиговка» (древесно-кустарниковые заросли), Пятихатки (древесно-кустарниковые заросли) (Рис. 17д, 18з, и).
Cydia fagiglandana (Zeller, 1841) – обычный лесной мезофил, Широкий олигофаг, дендрофаг (Fagaceae: Quercus sp., Fagus sp.), вредитель леса. Время лёта: май – сентябрь. Точки сбора в Приазовском БГР: Ростов-на-Дону, Ботанический сад ЮФУ (древесно-кустарниковые заросли), УОХ ЮФУ «Недвиговка» (древесно-кустарниковые заросли) (Рис. 17е, 18к).
Archips rosana (Linnaeus, 1758) – обычный лесной мезофил. Широкий олигофаг, дендрофаг (Rosaceae, Grossulariaceae, Juglandaceae, Caprifoliaceae, Adoxaceae, Betulaceae, Ericaceae, Oleaceae, Ranunculaceae, Salicaceae, Ulmaceae, Cornaceae, Elaeagnaceae, Fagaceae, Sapindaceae, Lythraceae, Rutaceae, Myricaceae, Fabaceae, Urticaceae, Apiaceae, Solanaceae, Pinaceae), вредитель ягодных культур (многоядный вредитель). Время лёта: май – сентябрь. Точки сбора в Азово-Егорлыкском БГР: Гирловский (водно-болотные угодья), Гигант (агроценоз, сады в населённом пункте), Зерноград (урбоценоз, сквер), Победа (агроценоз, сады в населённом пункте). Точки сбора в Приазовском БГР: Ростов-на-Дону, Ботанический сад ЮФУ (древесно-кустарниковые заросли), Таганрог (частный сектор), Реконструктор (фруктовый сад), Русский Колодец, (фруктовый сад), УОХ ЮФУ «Недвиговка» (древесно-кустарниковые заросли), Синявское (балка реки Донской Чулек) (разнотравная степь) (Рис. 17ж, 18л, м).
Сводные данные о выявлении новых и хозяйственно значимых видов листовёрток исследованных БГР. Для подведения итогов и понятной визуализации таких показателей, как общее количество видов, присутствие новых и/или вредоносных видов в двух и более ботанико-географических районах, удобно использовать сводные таблицы. Таблица 3 позволяет сводить важные сравниваемые признаки в одном инструменте, который удобно использовать для анализа и сопоставления рассматриваемых параметров в структуре тортрицидофаун исследованных БГР.
Таблица 3 – Сводная таблица данных о выявлении новых и хозяйственно значимых видов листовёрток исследованных БГР
| № п/п | Виды | АЕ | П | Новые АЕ | Новые П | Вредители АЕ | Вредители П |
|---|---|---|---|---|---|---|---|
| 1 | Phtheochroa pulvillana | - | + | - | - | - | - |
| 2 | Phtheochroa farinosana | + | - | - | - | - | - |
| 3 | Phtheochroa fulvicinctana | + | + | - | - | - | - |
| 4 | Cochylimorpha woliniana | - | + | - | Н | - | - |
| 5 | Cochylimorpha obliquana | - | + | - | - | - | - |
| 6 | Cochylimorpha subwoliniana | - | + | - | Н | - | - |
| 7 | Diceratura roseofasciana | - | + | - | - | - | - |
| 8 | Phalonidia albipalpana | + | + | - | - | - | - |
| 9 | Phalonidia manniana | + | - | - | - | - | - |
| 10 | Phalonidia contractana | + | + | - | - | - | - |
| 11 | Agapeta hamana | + | + | - | - | - | - |
| 12 | Eugnosta lathoniana | - | + | - | - | - | - |
| 13 | Eugnosta magnificana | + | + | - | - | - | - |
| 14 | Aethes williana | + | - | Н | - | - | - |
| 15 | Aethes nefandana | + | - | Н | - | - | - |
| 16 | Aethes tesserana | + | - | - | - | - | - |
| 17 | Aethes beatricella | - | + | - | - | - | - |
| 18 | Aethes bilbaensis | - | + | - | - | - | - |
| 19 | Cochylidia implicitana | - | + | - | Н | - | - |
| 20 | Neocochylis hybridella | - | + | - | - | - | - |
| 21 | Brevicornutia pallidana | + | + | - | - | - | - |
| 22 | Pontoturania posterana | + | + | - | - | - | - |
| 23 | Tortrix viridana | + | + | - | - | ВЛ | ВЛ |
| 24 | Aleimma loeflingiana | + | + | - | - | ВЛ | ВЛ |
| 25 | Acleris holmiana | - | + | - | - | - | ВПК |
| 26 | Acleris forsskaleana | - | + | - | - | - | - |
| 27 | Acleris rhombana | - | + | - | - | - | - |
| 28 | Acleris variegana | - | + | - | - | - | МВ |
| 29 | Acleris aspersana | + | - | - | - | ВПК | - |
| 30 | Acleris kochiella | - | + | - | - | - | ВЛ |
| 31 | Neosphaleroptera nubilana | - | + | - | - | - | ВПК |
| 32 | Doloploca punctulana | - | + | - | - | - | - |
| 33 | Eana incanana | - | + | - | - | - | - |
| 34 | Cnephasia incertana | + | + | - | - | - | - |
| 35 | Cnephasia stephensiana | + | + | - | - | ВДК | ВДК |
| 36 | Cnephasia alticolana | - | + | - | - | - | - |
| 37 | Cnephasia asseclana | - | + | - | - | - | ПВДК |
| 38 | Cnephasia pasiuana | - | + | - | - | - | ВЗЗК |
| 39 | Cnephasia genitalana | + | + | - | - | - | - |
| 40 | Cnephasia communana | + | + | - | - | - | - |
| 41 | Epagoge grotiana | - | + | - | Н | - | МВ |
| 42 | Paramesia gnomana | - | + | - | - | - | - |
| 43 | Periclepsis cinctana | + | - | - | - | - | - |
| 44 | Archips podana | + | + | - | - | ВПК | ВПК |
| 45 | Archips crataegana | - | + | - | - | - | ВПК |
| 46 | Archips xylosteana | + | + | - | - | ВПК | ВПК |
| 47 | Archips rosana | + | + | - | - | ВПК | ВПК |
| 48 | Choristoneura diversana | - | + | - | - | - | ВПК |
| 49 | Choristoneura hebenstreitella | - | + | - | - | - | МВ |
| 50 | Choristoneura lafauryana | - | + | - | - | - | - |
| 51 | Argyrotaenia ljungiana | + | + | - | - | - | - |
| 52 | Ptycholoma lecheana | - | + | - | - | - | МВ |
| 53 | Pandemis dumetana | - | + | - | Н | - | МВ |
| 54 | Aphelia ferugana | - | + | - | - | - | - |
| 55 | Clepsis spectrana | + | + | - | - | МВ | МВ |
| 56 | Clepsis pallidana | + | + | - | - | - | - |
| 57 | Clepsis neglectana | + | + | - | - | - | - |
| 58 | Lozotaeniodes formosana | + | - | - | - | ВЛ | - |
| 59 | Bactra lancealana | + | + | Н | Н | МВ | МВ |
| 60 | Bactra lacteana | + | - | Н | - | - | - |
| 61 | Endothenia gentianaeana | - | + | - | - | - | - |
| 62 | Endothenia marginana | - | + | - | - | - | - |
| 63 | Endothenia pullana | - | + | - | Н | - | - |
| 64 | Endothenia nigricostana | - | + | - | Н | - | - |
| 65 | Endothenia quadrimaculana | - | + | - | - | - | - |
| 66 | Hedya salicella | - | + | - | - | - | ВЛ |
| 67 | Hedya nubiferana | + | + | - | - | ВПК | ВПК |
| 68 | Hedya pruniana | + | + | - | - | МВ | МВ |
| 69 | Hedya ochroleucana | - | + | - | - | - | ВПК |
| 70 | Celypha striana | + | + | - | - | - | - |
| 71 | Celypha rosaceana | + | + | - | - | - | - |
| 72 | Celypha flavipalpana | - | + | - | - | - | - |
| 73 | Celypha cespitana | + | + | - | - | - | - |
| 74 | Celypha doubledayana | + | - | Н | - | - | - |
| 75 | Olethreutes arcuella | + | + | - | - | - | - |
| 76 | Lobesiodes euphorbiana | - | + | - | - | - | - |
| 77 | Lobesia botrana | + | + | - | - | МВ | МВ |
| 78 | Lobesia abscisana | + | + | - | - | - | - |
| 79 | Lobesia artemisiana | + | - | - | - | - | - |
| 80 | Thiodia citrana | + | + | - | - | - | - |
| 81 | Thiodia irinae | - | + | - | - | - | - |
| 82 | Thiodia torridana | - | + | - | - | - | - |
| 83 | Spilonota ocellana | - | + | - | - | - | ВЛ |
| 84 | Spilonota laricana | - | + | - | - | - | - |
| 85 | Epinotia tenerana | + | - | Н | - | ВЛ | - |
| 86 | Epinotia festivana | - | + | - | - | - | ВЛ |
| 87 | Zeiraphera ratzeburgiana | - | + | - | - | - | ВЛ |
| 88 | Zeiraphera isertana | - | + | - | - | - | МВ |
| 89 | Pelochrista fusculana | + | - | Н | - | - | - |
| 90 | Pelochrista metria | + | - | Н | - | - | - |
| 91 | Pelochrista caecimaculana | + | + | - | - | - | - |
| 92 | Pelochrista mollitana | - | + | - | Н | - | - |
| 93 | Pelochrista subtiliana | + | + | - | - | - | - |
| 94 | Pelochrista infidana | - | + | - | - | - | - |
| 95 | Pelochrista arabescana | + | - | - | - | - | - |
| 96 | Pelochrista labyrinthicana | + | - | - | - | - | - |
| 97 | Eucosma cana | + | + | - | - | - | - |
| 98 | Eucosma flavispecula | - | + | - | Н | - | - |
| 99 | Eucosma aemulana | - | + | - | - | - | - |
| 100 | Eucosma albidulana | - | + | - | - | - | - |
| 101 | Eucosma metzneriana | - | + | - | - | - | - |
| 102 | Eucosma tundrana | - | + | - | - | - | - |
| 103 | Eucosma wimmerana | - | + | - | Н | - | - |
| 104 | Eucosma conterminana | + | + | - | - | ВОК | ВОК |
| 105 | Eucosma pupillana | + | + | - | - | - | - |
| 106 | Eucosma luciana | + | - | - | - | - | - |
| 107 | Epibactra immundana | + | - | - | - | - | - |
| 108 | Gypsonoma minutana | + | + | - | - | ВЛ | ВЛ |
| 109 | Gypsonoma sociana | - | + | - | Н | - | ВЛ |
| 110 | Epiblema sticticana | + | + | - | - | - | - |
| 111 | Epiblema scutulana | + | + | - | - | - | - |
| 112 | Epiblema cirsiana | + | + | - | - | - | - |
| 113 | Epiblema foenella | - | + | - | - | - | - |
| 114 | Epiblema costipunctana | + | + | - | - | - | - |
| 115 | Epiblema graphana | + | + | - | - | - | - |
| 116 | Epiblema sarmatana | + | + | - | - | - | - |
| 117 | Notocelia cynosbatella | - | + | - | Н | - | - |
| 118 | Notocelia tetragonana | - | + | - | - | - | - |
| 119 | Notocelia uddmanniana | - | + | - | - | - | ВЯК |
| 120 | Notocelia roborana | - | + | - | - | - | ВДК |
| 121 | Notocelia trimaculana | - | + | - | Н | - | ВПК |
| 122 | Rhyacionia buoliana | - | + | - | - | - | ВЛ |
| 123 | Rhyacionia pinivorana | + | - | - | - | ВЛ | - |
| 124 | Enarmonia formosana | + | - | Н | - | ВПК | - |
| 125 | Ancylis comptana | - | + | - | - | - | - |
| 126 | Ancylis unculana | + | + | - | - | ВЛ | ВЛ |
| 127 | Ancylis apicella | - | + | - | - | - | ВЛ |
| 128 | Ancylis achatana | - | + | - | - | - | ВПК |
| 129 | Ancylis mitterbacheriana | + | + | - | - | МВ | МВ |
| 130 | Aspila funebrana | + | + | - | - | ВПКК | ВПКК |
| 131 | Aspila janthinana | - | + | - | - | - | ВПКК |
| 132 | Aspila molesta | + | + | - | - | ВПК | ВПК |
| 133 | Grapholita compositella | - | + | - | - | - | ВСК |
| 134 | Grapholita delineana | - | + | - | - | - | - |
| 135 | Grapholita coronillana | - | + | - | - | - | - |
| 136 | Grapholita caecana | - | + | - | - | - | - |
| 137 | Ephippiphora gemmiferana | - | + | - | - | - | - |
| 138 | Cydia nigricana | - | + | - | - | - | ВЗБК |
| 139 | Cydia pomonella | + | + | - | - | ВПК | ВПК |
| 140 | Cydia pyrivora | + | + | - | - | ВПСК | ВПСК |
| 141 | Cydia servillana | - | + | - | Н | - | - |
| 142 | Cydia splendana | + | + | - | - | ВЛ | ВЛ |
| 143 | Cydia fagiglandana | + | + | - | - | ВЛ | ВЛ |
| 144 | Cydia amplana | + | + | - | - | ВЛ | ВЛ |
| 145 | Pammene suspectana | + | + | - | - | - | - |
| 146 | Dichrorampha simpliciana | - | + | - | - | - | - |
| 147 | Dichrorampha incognitana | - | + | - | - | - | - |
| 148 | Dichrorampha plumbana | - | + | - | Н | - | - |
| 149 | Dichrorampha vancouverana | + | - | Н | - | - | - |
| 150 | Dichrorampha agilana | + | + | Н | Н | - | - |
Примечания к табл. 3: «+» – присутствие; «-» – отсутствие; Н – новый для Ростовской области вид; ВЛ – вредитель леса; ВПК – вредитель плодовых культур; МВ – многоядный вредитель; ВДК – вредитель декоративных культур; ВПКК – вредитель плодовых косточковых культур; ВЯК – вредитель ягодных культур; ВЗБК – вредитель зерновых бобовых культур; ВЗЗК – вредитель зерновых злаковых культур; ВОК – вредитель овощных культур; ВПСК – вредитель плодовых семечковых культур; ПВДК – потенциальный вредитель декоративных культур.
Распределение листовёрток по основным типам местообитаний и анализ сходства их сообществ в исследованных БГР. Для оценки сходства сообществ листовёрток в типизированных местообитаниях (Табл. 4 и 5) были построены соответствующие дендрограммы (Рис. 19).
Таблица 4 – Список местообитаний Азово-Егорлыкского БГР с указанием условных номеров
| № | Основные типы местообитаний | ΣS | ΣN |
|---|---|---|---|
| 1 | Водно-болотные угодья | 1 | 5 |
| 2 | Древесно-кустарниковые заросли | 10 | 187 |
| 3 | Древесно-кустарниковые насаждения | 1 | 1 |
| 4 | Плодово-ягодные сады | 72 | 710 |
| 5 | Скверы в плотной городской застройке | 6 | 10 |
| 6 | Разнотравные степи | 1 | 1 |
Таблица 5 – Список местообитаний Приазовского БГР с указанием условных номеров
| № | Основные типы местообитаний | ΣS | ΣN |
|---|---|---|---|
| 1 | Байрачные и низинные леса | 11 | 39 |
| 2 | Плодово-ягодные сады | 12 | 108 |
| 3 | Древесно-кустарниковые заросли | 124 | 4331 |
| 4 | Древесно-кустарниковые насаждения | 5 | 1274 |
| 5 | Частные секторы | 1 | 7 |
| 6 | Остепнённые балочные склоны и овраги | 6 | 10 |
| 7 | Закустаренные остепнённые склоны и овраги | 9 | 24 |
| 8 | Разнотравные степи | 6 | 10 |
| 9 | Степные залежи | 1 | 1 |
| 10 | Ковыльные степи | 3 | 3 |
В Азово-Егорлыкском БГР максимальный уровень сходства сообществ наблюдается между древесно-кустарниковыми зарослями (2) и плодово-ягодными садами (4). Наиболее специфичным по биоразнообразию по сравнению со всеми остальными типами местообитаний выступают разнотравные степи (6) (Рис. 19а).
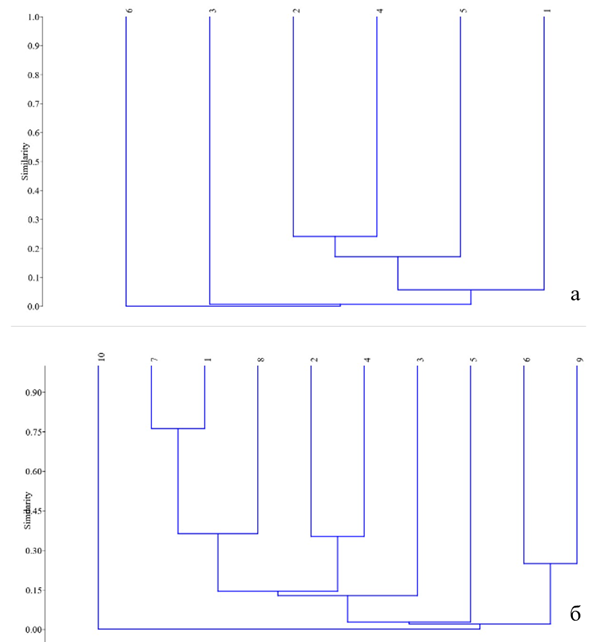
Рис. 19 – Дендрограмма сходства сообществ листовёрток: а –Азово-Егорлыкский БГР; б – Приазовский БГР (коэффициент Чекановского-Съёренсена, алгоритм UPGMA в PAST3)
В Приазовском БГР самая высокая степень сходства (0,75) характерна для группы, объединяющей байрачные и низинные леса, а также закустаренные остепнённые склоны и овраги. Эта же группа демонстрирует заметное сходство (0,5) с разнотравными степями. Высокий уровень сходства также отмечен между остепнёнными балочными склонами и оврагами (6) и степными залежами (9). Наиболее обособленным местообитанием в данном БГР являются ковыльные степи (10) (Рис. 19б).
Анализ биологического разнообразия. На территории Южных Приазовских степей максимальное значение индекса Шеннона зафиксировано для плодово-ягодных садов (H=2,73), тогда как скверы в плотной городской застройке значительно уступают им (H=1,5). Индекс Симпсона (D) подтверждает это распределение: плодово-ягодные сады – D=0,85, скверы – D=0,7 (Рис. 20).
Графическое представление индекса Бриллуена (HB) также демонстрирует преобладание плодово-ягодных садов (HB=2,58); субдоминантой выступают древесно-кустарниковые заросли (HB=1,0). Следует отметить, что местообитания с нулевым значением индекса являются наиболее однородными, поскольку содержат лишь по одному экземпляру одного вида. Такая ситуация характерна для водно-болотных угодий, древесно-кустарниковых насаждений и разнотравных степей.
Обратная зависимость между величиной индекса и биоразнообразием наблюдается в случае индекса Бергера-Паркера (d), который отражает степень доминирования одного вида в пределах местообитания. Наименьшее значение (d=0,32) зафиксировано для плодово-ягодных садов, где видовое разнообразие наиболее высоко. Значение, равное единице, характерно для местообитаний, с которых было собрано несколько экземпляров одного вида (водно-болотные угодья, древесно-кустарниковые насаждения, разнотравные степи); в таких случаях данный вид становится явной доминантой ввиду отсутствия альтернатив.
Для территории Приазовского БГР использованы те же аналитические подходы. Согласно индексам Симпсона и Шеннона, частные секторы и степные залежи имеют нулевые значения. Самое высокое видовое разнообразие отмечено для древесно-кустарниковых зарослей (D=0,91, H=2,965) и закустаренных остепнённых склонов и оврагов (D=0,86, H=2,14), что свидетельствует о значительном числе видов и собранных экземпляров. Индекс Бриллуена подтверждает это распределение: доминанта – древесно-кустарниковые заросли (HB=2,9), субдоминанта – закустаренные остепнённые склоны и овраги (HB=1,7) (Рис. 20).
Как отмечалось выше, индекс Бергера-Паркера отражает степень доминирования видов в сообществе. На севере Приазовья наименьшие значения индекса зафиксированы для древесно-кустарниковых зарослей (d=0,165), закустаренных остепнённых склонов и оврагов (d=0,2), а также для плодово-ягодных садов (d=0,61) и остепнённых балочных склонов и оврагов (d = 0,3). Наибольшие значения индекса характерны для частных секторов и степных залежей, что указывает на крайнюю скудность сборов на этих местообитаниях (по одному экземпляру одного вида) и, следовательно, их очевидное доминирование.
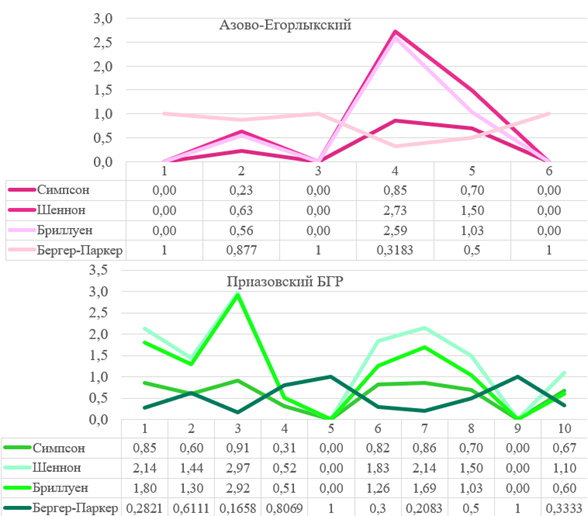
Рис. 20 – Индексы биоразнообразия и доминирования исследуемых БГР
На основании полученных данных можно заключить, что значения индексов биоразнообразия для Северных Приазовских степей в целом выше, чем для Южных, что свидетельствует о более высокой степени изученности Приазовского БГР.
Расчёт и анализ потенциально возможного числа видов. Для проверки данного утверждения, учёта вырожденности местообитаний по сборам, а также для определения наиболее перспективных (с точки зрения выявления новых для региона видов) точек для экспедиционных выездов применяется метод разрежения и расчёта потенциально возможного числа видов с использованием эстиматоров (Сушко, 2021), в частности – кривых ассамблей (Рис. 21).
Для Южных Приазовских степей характерны достаточно плавные кривые, приближающиеся к асимптоте: например, для плодово-ягодных садов (4) и древесно-кустарниковых зарослей (2). Это указывает на то, что фактическое видовое богатство этих местообитаний может быть значительно выше выявленного (Рис. 21а).
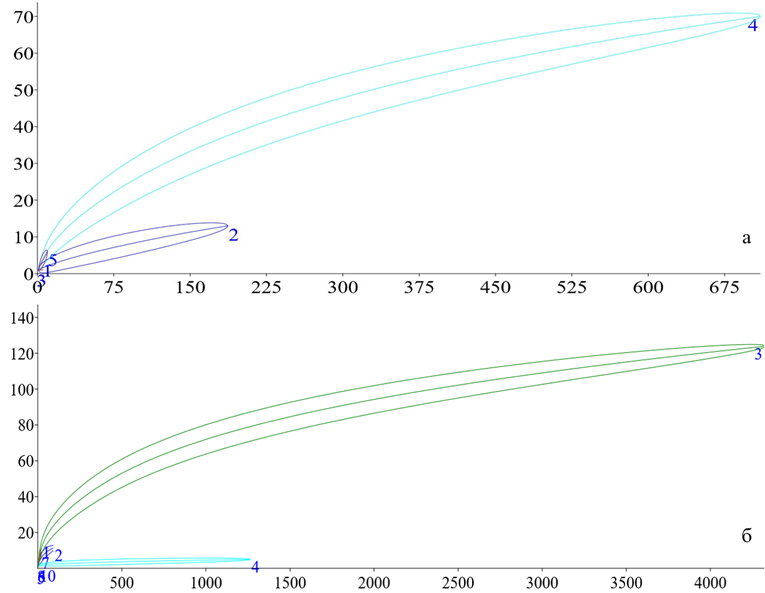
Рис. 21 – Кривые разрежения ассамблей тортрицид: а – Азово-Егорлыкский БГР (доминанты: 4 – плодово-ягодные сады, 2 – древесно-кустарниковые заросли); б – Приазовский БГР (доминанты: 3 – древесно-кустарниковые заросли, 4 – древесно-кустарниковые насаждения)
В случае Северных Приазовских степей кривая 4 (древесно-кустарниковые насаждения) имеет вид асимптоты: после пятого экземпляра она выходит на плато, что свидетельствует о том, что обнаруженное число видов практически соответствует потенциально возможному для данного типа местообитаний. Кривая 3 (древесно-кустарниковые заросли), напротив, повторяет ситуацию, описанную для Южных степей, и указывает на возможность получения гораздо большего разнообразия, чем уже установлено (Рис. 21б).
Календарь-схема плотности сборов листовёрток исследованных БГР. Основываясь на календаре-схеме плотности сборов (Рис. 22), можно сделать выводы об интенсивности исследований листовёрток в Приазовском и Азово-Егорлыкском ботанико-географических районах.
На календаре визуально доминирует Приазовский БГР (зелёный цвет), особенно в периоды максимальной активности. Это свидетельствует о том, что эта территория исследована значительно интенсивнее и имеет более высокую степень «обловленности» по сравнению с Азово-Егорлыкским БГР (розовый цвет) на протяжении большей части сезона.
Общую интенсивность исследований можно представить в виде следующей сезонной структуры:
Март – начало сезона:
- Сборы в марте (1–2 даты) отмечены только в Приазовском БГР, ранневесенние исследования проводились исключительно в этом районе. Азово-Егорлыкский БГР в марте не обследовался.
Апрель – нарастание активности:
- В апреле (3–4 даты) представлены оба района, но доминирует Приазовский. Розовый цвет присутствует, но в меньшей степени. Это указывает на то, что оба района уже активно изучаются, но Приазовский остается в приоритете.
Май – первый пик активности:
- В мае наблюдается максимальное доминирование зелёного цвета (7–9 дат). Основная масса сборов в период наивысшей активности приходится на Приазовский БГР.
- Азово-Егорлыкский БГР в мае представлен слабо, что говорит о его соподчиненной роли в этот ключевой период.
Июнь – период высокой активности:
- В июне ситуация выравнивается (5–6 дат). Здесь розовый и зелёный представлены практически поровну. Это единственный месяц, когда интенсивность сборов в Азово-Егорлыкском БГР сравнялась с Приазовским.
Июль – спад активности:
- В июле оба района представлены примерно одинаково (3–4 даты), но общая интенсивность низкая. В этот период на обеих территориях проводились локальные или дополнительные/поддерживающие сборы.
Август – второй пик активности:
- В августе снова наблюдается перевес в сторону Приазовского БГР (5–6 дат). Исследователи вновь сместили фокус на этот район.
Сентябрь – затухание активности и завершение сезона:
- В сентябре оба района присутствуют примерно равномерно (3–4 даты), завершая полевой сезон.
Таким образом, Приазовский БГР является гораздо более изученной территорией. Именно на него приходится пик активности в мае, ранневесенние сборы в марте и второй пик активности в августе. Азово-Егорлыкский БГР изучался менее интенсивно и скорее эпизодически. Его роль возрастает лишь в июне, когда интенсивность исследований сравнялась с Приазовским БГР. В остальные месяцы (особенно в мае) он значительно уступает Северным Приазовским степям по количеству сборов (Рис. 22). Это связано как с логистической доступностью территорий, так и с изначальными научными приоритетами исследователей, сперва сосредоточивших основные усилия на Приазовском БГР (см. Слюнькова и др., 2025).

Рис. 22 – Календарь-схема плотности сборов листовёрток исследованных БГР
Заключение
В Приазовском ботанико-географическом районе (данные за 1937–2025 гг.) выявлено 128 видов листовёрток (5812 экз.), в Азово-Егорлыкском (–2025 гг) – 75 видов (914 экз.). В Приазовском БГР к подсемейству Tortricinae относится 50 видов (40%), к Olethreutinae – 78 (60%); в Азово-Егорлыкском – 28 (37%) и 47 (63%) соответственно. В обоих районах доминируют «очень редкие» виды (68 в Северных Приазовских степях, 39 в Южных), что связано с регулярными единичными находками новых для региона таксонов. Соотношение «обычных» (28 и 20) и «локальных» (14 и 10) видов сходно, тогда как редких видов в Приазовском БГР в 1,7 раза больше (17 против 10).
Распределение по экологическим группам обнаруживает сходную картину: повсеместно доминируют лесные мезофилы (49 видов в Северных степях, 24 – в Южных), субдоминантами выступают лугово-лесные мезофилы (22 и 11). Это свидетельствует о формировании лесо-лугового мезофильного экологического субкомплекса тортрицид в исследованных БГР.
Трофический спектр обоих районов помимо Asteraceae и Rosaceae включает Fabaceae, Fagaceae, Betulaceae и Ericaceae. В Южных Приазовских степях четвёртое место занимает Betulaceae, в Северных – Fagaceae. Общее ядро составляют хортофаги (38 видов в Южных степях, 64 – в Северных), дендрофаги (17 и 28) и дендро-тамнофаги (7 и 13). В Северных степях отмечено большее разнообразие «переходных» групп, что объясняется более сложной структурой растительных сообществ.
Ведущую группу в обоих БГР образуют виды, не имеющие хозяйственного значения (47 в Южных, 75 в Северных), многие из которых являются хортофагами на дикорастущих травах. Ядро хозяйственно значимых видов представлено вредителями леса (по 16 видов в каждом районе), плодовых культур (по 13) и многоядными вредителями (по 12).
Наиболее высокий уровень сходства сообществ в Азово-Егорлыкском БГР отмечен для древесно-кустарниковых зарослей и плодово-ягодных садов; в Приазовском БГР – для байрачных и низинных лесов, а также закустаренных остепнённых склонов и оврагов. В Южных Приазовских степях наиболее разнообразны плодово-ягодные сады (H=2,73; D=0,85; HB=2,58; индекс Бергера–Паркера – 0,32); скверы в городской застройке значительно уступают (H=1,5; D=0,7). В Северных степях лидируют древесно-кустарниковые заросли (D=0,91; H=2,965; индекс Бергера–Паркера – 0,165) и закустаренные остепнённые склоны с оврагами (D=0,86; H=2,14; индекс Бергера–Паркера – 0,2).
Кривые накопления видов для плодово-ягодных садов и древесно-кустарниковых зарослей в Азово-Егорлыкском БГР имеют плавную форму, не достигающую асимптоты, что предполагает более высокое потенциальное видовое богатство. В Северных Приазовских степях кривая для древесно-кустарниковых насаждений достигает асимптоты, указывая на практически полную выявленность видового состава, тогда как кривая для зарослей асимптоты не достигает, повторяя ситуацию Южных степей.
Литература/References
1) Алфераки С.Н. Чешуекрылые окрестностей Таганрога // Труды Русского энтомологического общества. – 1876. – Т. 8, № 2–3. – С. 150–226.
2) Алфераки С.Н. Чешуекрылые окрестностей Таганрога // Труды русского энтомологического общества. – 1877. – Т. 10, № 1. – С. 35–53.
3) Алфераки С.Н. Чешуекрылые окрестностей Таганрога // Труды Русского энтомологического общества. – 1880. – Т. 11. – С. 45–50.
4) Алфераки С.Н. Чешуекрылые окрестностей Таганрога // Труды русского энтомологического общества. – 1908. – Т. 38. – С. 558–618.
5) Артохин К.С., Полтавский А.Н. Злаковая листовёртка в Ростовской области // Защита и карантин растений. – 2018. – № 8. – С. 28–30.
6) Артохин К.С., Полтавский А.Н. Злаковая листовёртка на юге России // Защита и карантин растений. – 2021. – № 1. – С. 25–28.
7) Архангельский Н.Н. Луговой мотылёк и меры борьбы с ним // Краевая станция защиты растений. Серия Биологическая. – 1929. – № 19. – С. 1–8.
8) Архангельский Н.Н. Огородные вредители. – Ростов-на-Дону: Сев. Кавказ, 1931. – 47 с.
9) Данилевский А.С., Кузнецов В.И. Листовёртки (Tortricidae), Триба Плодожорки Laspeyresiini // Фауна СССР. Насекомые чешуекрылые. – Л.: Наука, 1968. – Т. 5. – Вып. 1. – 635 с. 78
10) Добровольский Б.В. Луговой мотылек (в условиях Нижнего Дона и Предкавказья). – Ростов-на-Дону: Ростиздат, 1939. – 124 с.
11) Добровольский Б.В. Вредители полевых культур (на Дону). – Ростовна-Дону: Ростиздат, 1950. – 132 с.
12) Добровольский Б.В. Распространение вредных насекомых: Очаги и зоны наибольшей вредности. – Москва: Сов. наука, 1959. – 215 с.
13) Дунаев Е.А. Методы эколого-энтомологических исследований. – Москва: МосгорСЮН, 1997. – 44 с.
14) Ершов В., Фильд А. Каталог чешуекрылых Российской Империи // Труды Русского энтомологического общества. – 1870. – Т. 4. – С.130–204.
15) Загуляев А.К., Кузнецов В.И., Стекольников А.А., Сухарева И.Л., Фалькович М.И. Определитель насекомых европейской части СССР. Том IV. Чешуекрылые. Первая часть. – Ленинград: Наука, 1978. – 712 с.
16) Зверезомб-Зумбовский Е.В. Краткий отчёт о деятельности Донского бюро по борьбе с вредителями с-х растений в 1917 году. – Ростов н/Д, 1918. – 36 c.
17) Зверезомб-Зумбовский Е.В. Отчёт о деятельности Донского бюро по борьбе с вредителями с-х растений за 1918 год. – Ростов н/Д, 1919. – 12 с.
18) Зозулин Г.М. Пашков Г.Д. Геоботаническое районирование: Нижний Дон (Ростовская область) // Растительные ресурсы. – Ростов-на-Дону [б. и.], 1980. – Часть 1. Леса. – С. 40–48.
19) Корнелио М.П. Школьный атлас-определитель бабочек. – Москва: Просвещение, 1986. – 255 с.
20) Косенко Т.Г. Зональные особенности сельскохозяйственного производства Ростовской области // Международный научно-исследовательский журнал, 2023. – № 1 (127). – С. 2–4.
21) Недошивина С.В. Листовёртки трибы Olethreutini (Lepidoptera, Tortricidae) фауны России: автореферат дис. ... канд. биол. наук: 03.02.05. – Санкт-Петербург, 2010. – 19 с.
22) Недошивина С.В. Экологические особенности листовёрток (Lepidoptera, Tortricidae) трибы Olethreutini фауны России // Энтомологические и паразитологические исследования в Поволжье. – 2011. – № 9. – С. 68–77.
23) Определитель насекомых Дальнего Востока России. Т. V. Ручейники и чешуекрылые. – Владивосток: Дальнаука, 2001. – Часть 3. – 621 с.
24) Определитель насекомых юга России. – Ростов-на-Дону: Foundation, 2016. – 1050 с.
25) Пестициды.ru. – URL: https://www.pesticidy.ru (дата обращения: 20.02.2026).
26) Полтавский А.Н., Артохин К.С. Энтомологические рефугиумы и их значение при ведении Красной книги Ростовской области. – Ростов-на-Дону: Изд-во «ИП Кубеш», 2012. – 184 с.
27) Полтавский А.Н., Колесник О.А. Листовёртки (Lepidoptera: Tortricidae), вредящие древесным растениям ботанического сада ЮФУ // Роль ботанических садов в сохранении и мониторинге биоразнообразия: Международная научная конференция, посвященная 100-летию Южного федерального университета, Ростов-на-Дону, 27–30 мая 2015 г. – Ростов-на-Дону: Изд-во ЮФУ, 2015. – С. 80– 83.
28) Померанцев Д.В. Вредные насекомые и борьба с ними в лесах и лесных полосах юго-востока Европейской части СССР. – Москва; Ленинград: Гослесбумиздат, 1949. – 212 с.
29) Романова В.П. Листовёртки и их паразиты на древесных породах в окрестностях г. Ростова-на-Дону // Учёные записки Ростовского государственного университета. – 1949. – Т. 15, Вып. 6. – С. 47–61.
30) Романова В.П. Вредные насекомые ботанического сада // Сборник трудов Ботанического сада РГУ. – 1956. – Т. 35, Вып. 2. – С. 77–87.
31) Романчук Р.В., Кладковая О.А. Экофаунистический обзор листовёрток (Lepidoptera: Tortricidae) охраняемого ландшафта «Фоминская дача» Ростовской области. Часть 1: таксономический состав, обилие, распределение по экологическим группам и местообитаниям // Актуальные проблемы экологии и природопользования: сборник материалов. – Ростов-на-Дону; Таганрог: Изд-во ЮФУ, 2021а. – С. 111–116.
32) Романчук Р.В., Кладковая О.А. Экофаунистический обзор листовёрток (Lepidoptera: Tortricidae) охраняемого ландшафта «Фоминская дача» Ростовской области. Часть 2: трофическая приуроченность, широта трофических связей, хозяйственное значение // Актуальные проблемы экологии и природопользования: сборник материалов. – Ростов-на-Дону; Таганрог: Изд-во ЮФУ, 2021б. – С. 117–121.
33) Романчук Р.В., Кладковая О.А. Каталог коллекции животных кафедры зоологии Академии биологии и биотехнологии им. Д.И. Ивановского Южного федерального университета. Сообщение 2. Отряд Чешуекрылые – Lepidoptera, семейство листовёртки – Tortricidae // Ученые записки Крымского федерального университета имени В.И. Вернадского Биология. Химия. – 2022. – Том 8 (74), № 1. – С. 140–155. 81
34) Романчук Р.В., Колесников С.И. Эколого-фаунистическая характеристика чешуекрылых (Insecta: Lepidoptera) особо охраняемых природных территорий Ростовской области: монография. – Ростов-на-Дону; Таганрог: Издательство Южного федерального университета, 2022. – 368 с. – DOI: 10.18522/801299932
35) Романчук Р.В., Елфимова Н.С., Пивоварова Д.Д., Кладковая О.А., Минникова Т.В. Чешуекрылые насекомые (Insecta: Lepidoptera) ООПТ ГПЗ «Горненский» (Ростовская область): эколого-фаунистический обзор. – 2023а. – Свидетельство о государственной регистрации базы данных № 2023623278 от 28.09.2023 г.
36) Романчук Р.В., Кладковая О.А., Дмитриева А.А. Листовёртки (Lepidoptera, Tortricidae) ООПТ «Фоминская дача» (Ростовская область): экологофаунистический обзор. – 2023б. – Свидетельство о государственной регистрации базы данных № 2023620071 от 23.12.2022.
37) Романчук Р.В., Мищенко Д.П., Слюнькова В.Г., Гузь А.А. Бабочки листовёртки Южноевропейской Lepidoptera: Tortricidae) Верхнедонского района Ростовской области: аннотированный список с указанием хозяйственно значимых видов // Воспроизводство, использование, охрана, защита и биоразнообразие лесов на юге европейской части России: сборник статей, посвященный 75-летию Южноевропейской научно-исследовательской лесной опытной. – Пушкино: ВНИИЛМ, 2024а. – С. 256–260.
38) Романчук Р.В., Слюнькова В.Г., Мищенко Д.П., Гузь А.А., Елфимова Н.С., Мижерицкая А.Р. Хозяйственно значимые таксоны листовёрток (Lepidoptera: Tortricidae) Ростовской области: фаунистический состав, данные о 82 выявлении и распространении в регионе (предварительный анализ) // Учёные записки Крымского федерального университета имени В.И. Вернадского. Биология. Химия. – 2024б. – Том 10 (76), № 3. – С. 162–190. – DOI: 10.29039/2413- 1725-2024-10-3-162-190
39) Синёв С.Ю. (Ред.). Каталог чешуекрылых (Lepidoptera) России. Издание 2-е. – Санкт-Петербург: Зоологический институт РАН, 2019. – 448 с.
40) Слюнькова В.Г., Мижерицкая А.Р., Романчук Р.В., Шидловская Л.Г., Святоха А.М. Сообщества листовёрток (Lepidoptera: Tortricidae) Приазовского ботанико-географического района (Ростовская область) // Живые и биокосные системы. – 2025. – №52. – URL: https://jbks.ru/archive/issue-52/article-10. – DOI: 10.18522/2308-9709-2025-52-10
41) Сушко Г.Г. Программное обеспечение PAST – как инструмент анализа данных в фаунистических исследованиях // Известия Гомельского государственного университета имени Ф. Скорины – 2021. – № 3 (126). – 54–61.
42) Фишкин М.В., Узденов А.М. Природно-заповедный фонд Тихого Дона. – Ростов-на-Дону: Изд-во «D&V», 2018. – 304 c.
43) Шайхутдинова А. А. Методы оценки биоразнообразия: Методические указания // Оренбургский гос. ун-т. – Оренбург: ОГУ, 2019. – 37 с.
44) Шестакова А.В. Яблонная плодожорка и меры борьбы с ней в Ростовской области: Автореф. дис… канд. биол. наук. – Ростов-на-Дону, 1980. – 22 с.
45) Шидловская Л.Г., Святоха А.М., Романчук Р.В., Слюнькова В.Г., Мижерицкая А.Р. Таксономическая организация и экологическое разнообразие сообществ листовёрток (Lepidoptera: Tortricidae) Долины Нижнего Дона // Живые и биокосные системы. – 2026. – № 55; URL: https://jbks.ru/archive/issue-55/article-10. – DOI: 10.18522/2308-9709-2026-55-10
46) EPPO Global Database. – URL: https://gd.eppo.int (дата обращения: 23.02.2026).
47) Lepiforum: Bestimmung von Schmetterlingen (Lepidoptera) und ihren Präimaginalstadien. – URL: http://www.lepiforum.de (дата обращения: 20.02.2026).
48) Moth Dissection. – URL: https://mothdissection.co.uk (дата обращения: 20.02.2026).
49) Poltavsky A.N. An Inventory of Tortricidae (Lepidoptera) from the Rostovon-Don province of Russia // Zootaxa. – 2015. – V. 4048, I. 4. – P. 538–552. – DOI: 10.11646/zootaxa.4048.4.5
50) Tortricid.net. Tortricidae Resources on the Web. – Version 2.0. – URL: http://www.tortricidae.com (дата обращения: 20.02.2026).
51) Zhang Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. – Zootaxa. – Auckland: Magnolia Press, 2011. – Vol. 3148. – P. 212–221.
Статья поступила в редакцию 18 февраля 2026 г.
Поступила после доработки 11 марта 2026 г.
Принята к печати 25 марта 2026 г.
Received 18, February, 2026
Revised 11, March, 2026
Accepted 25, March, 2026