1. Введение
Изучение синтеза нерибосомальных пептидов (НРП) началось в начале 1960-х годов с исследований биосинтеза циклических пептидов, содержащих D-аминокислоты, таких как тироцидин, производимый видами Bacillus. Татум и соавторы показали, что на биосинтез тироцидина в клетках не влияют ингибиторы рибосом, что привело к гипотезе, что его синтез происходит по механизму, отличному от трансляции белка (Mach, Reich, Tatum, 1963). В дальнейшем Фриц Липманн и Сёрен Лаланд продемонстрировали, что активация аминокислот осуществляется через образование тиоэфирных связей с 4′-фосфопантетеиновыми кофакторами в АТФ-зависимой реакции, сопровождающейся выделением пирофосфата и AMP. Ковалентное связывание аминокислот с ферментами открыло представление о роли Ppant-переносчика в каждом модуле НРПС, а пошаговый характер сборки — о модульной организации комплекса (Süssmuth, Mainz, 2017).
С конца XX века интенсивно развивались молекулярно-биологические методы: клонирование и экспрессия генов нерибосомальных пептидсинтаз/синтетаз (НРПС) in vitro, направленная инактивация отдельных компонентов, а затем полногеномное секвенирование микроорганизмов-продуцентов. В 1988 году группа Марахиеля расшифровала нуклеотидную последовательность гена tycA, кодирующего тироцидин-синтетазу, что стало первым генетическим подтверждением структуры НРПС (Weckermann, Fürbass, Marahiel, 1988). Уже через год ими был описан оперон биосинтеза грамицидина C, включающий гены grsA и grsB. Тем самым была окончательно подтверждена модульная организация этих многофункциональных энзимов, что заложило основу для предсказания структуры нерибосомальных пептидов по данным секвенирования (Krätzschmar, Krause, Marahiel, 1989).
Нерибосомальные пептидсинтетазы НРПС представляют собой мультидоменные ферментативные комплексы, которые катализируют синтез структурно разнообразных пептидов без участия рибосом и могут включать широкий спектр субстратов, включая стандартные и непротеиногенные аминокислоты, жирные кислоты, α-гидроксикислоты, α-кетокислоты и гетероциклы, что обеспечивает химическое и структурное разнообразие нерибосомальных пептидов (Stanišić, Kries, 2019). Сходный принцип также применяется бактериями для синтеза поликетидов с помощью поликетидсинтетаз (ПКС), только в их случае субстратом служат малонил-CoA и его производные. Некоторые биосинтетические кластеры содержат гибридные НРПС-ПКС системы, сочетающие оба механизма синтеза в единой ферментативной архитектуре.
Молекулярная масса НРПС может варьироваться в широких пределах. Так, ферменты, производящие небольшие пептиды имеют массу около 100 кДа, а крупнейшая из известных непрерывных бактериальных НРПС - синтетаза колоссина А - состоит из 15 модулей и 45 доменов, с молекулярной массой 1,8 МДа (Bode и др., 2015; Cai, Zhao, Bode, 2023; Iacovelli, Bovenberg, Driessen, 2021).
Хотя нерибосомальные пептиды продуцируются различными группами бактерий, основными филумами, ответственными за производство большинства бактериальных НРП, являются Proteobacteria, Actinobacteria, Firmicutes и Cyanobacteria (Borsetto и др., 2019; Wang и др., 2014). Накопление кластеров генов НРПС в этих четырёх филумах может частично отражать их сложный экологический образ жизни и потребность в механизмах химической защиты и коммуникации (Wang и др., 2014).
В рамках Proteobacteria ключевыми продуцентами выступают Gammaproteobacteria (особенно псевдомонады, синтезирующие сидерофоры и циклические липопептиды) и Deltaproteobacteria (преимущественно миксобактерии, обладающие разнообразным вторичным метаболизмом), тогда как Betaproteobacteria рассматриваются как потенциальный источник новых НРП (Baldim и др., 2017; Kooner, Soby, 2022; Léger и др., 2021). Actinobacteria, в частности стрептомицеты, демонстрируют высокую продуктивность благодаря крупным геномам, содержащим множественные кластеры НРПС и поликетидсинтетаз, что обуславливает синтез значительной доли известных антибиотиков и биоактивных соединений (Jian и др., 2023; Komaki и др., 2015; Sharma, Kaur, Salwan, 2021). Представители Firmicutes, такие как Bacillus, продуцируют структурно разнообразные пептиды, включая антибиотики (грамицидин, бацитрацин) и липопептиды (Kalyvas и др., 2025; Saiyam и др., 2024; Waongo и др., 2025). Cyanobacteria синтезируют биотоксичные НРП (например, микроцистины), играющие критическую роль в экологии водных систем (Hartnell и др., 2020). Таким образом, данные филумы представляют собой важнейшие источники НРП с широким спектром биологической активности и значительным фармакологическим потенциалом (Wang и др., 2014).
Благодаря своему структурному разнообразию и высокой биологической активности нерибосомальные пептиды нашли широкое применение в медицине и сельском хозяйстве. В медицинской сфере НРП применяются в качестве антибиотиков, иммунодепрессантов и противоопухолевых агентов. В сельском хозяйстве они используются как агенты биологического контроля (Agrawal и др., 2017; Felnagle и др., 2008; Kaspar, Neubauer, Gimpel, 2019; Ranjan и др., 2023). Изучаются возможности применения нерибосомально синтезируемых сидерофоров для селективного извлечения металлов из сложных промышленных растворов, что стало бы более экологичным и эффективным по сравнению с традиционными методами при низких концентрациях целевых катионов (Sieber и др., 2025).
Целью данного обзора является систематизация имеющейся информации о молекулярно-генетических механизмах нерибосомального синтеза у прокариот, определение малоизученных областей, перспектив развития данной области исследований и потенциально новых областей применения продуктов нерибосомального синтеза.
2. Механизм работы и строение НРПС
Синтез пептидов нерибосомальными пептидсинтетазами осуществляется посредством образования амидных связей между аминоацильными или другими ацильными мономерами. В отличие от рибосомального синтеза, где все аминокислоты добавляются на одном и том же активном центре, НРПС функционируют по модульному принципу: для взаимодействия с каждым аминокислотным остатком задействован отдельный набор ферментативных доменов. Совокупность таких доменов образует модуль. Выделяют 3 типа модулей: элонгации, инициации и терминации.
Модуль инициации, с которого начинается синтез пептида, как правило, включает домен аденилирования (A) и домен тиолирования (T), также известный как домен белка-переносчика пептидилов (PCP). За модулем инициации следуют один или несколько модулей элонгации, отличающихся наличием домена конденсации (С), расположенного перед доменами А и PCP. Синтез пептида начинается с активации субстрата в домене аденилирования, который может распознавать и активировать как протеиногенные, так и непротеиногенные аминокислоты, такие как D-формы протеиногенных аминокислот, орнитин, иминокислоты и т.д. (Iacovelli, Bovenberg, Driessen, 2021; Miller, Gulick, 2016; Walsh, O’Brien, Khosla, 2013). На данном этапе в присутствии Mg-АТФ А-домен аденилирует аминокислоту, образуя аминоацил-АМФ, который затем взаимодействует с тиоловой группой 4′-фосфопантетеина (PP) домена PCP, с образованием комплекса пептидил-PCP. Пептидил-PCP взаимодействует с донорным туннелем доменом конденсации следующего модуля, образуя комплекс PCP-C, в результате чего происходит активация А-домена этого модуля и описанный ранее процесс повторяется в этом модуле вплоть до момента образования пептидил-PCP, который взаимодействует сначала с акцепторным туннелем собственного С доменом, образуя комплекс PCP-С-PCP, C-домен катализирует образование пептида с образованием дипептидного промежуточного продукта, связанного с Т-доменом второго модуля и переносится в донорный туннель домена конденсации следующего модуля. Затем модуль инициации может загрузить другую аминокислоту субстрата и начать сборку следующего пептида (Corpuz, Sanlley, Burkart, 2022; Mansour, Gauld, 2024; Shi и др., 2020). Промежуточный пептидилтиоэфир передается от одного модуля к следующему, при этом в каждом модуле добавляется одна аминокислота. Цикл повторяется, пока пептид не достигнет модуля терминации, содержащего тиоэстеразный домен (TE), который либо катализирует реакцию гидролиза и высвобождает линейный пептид, либо катализирует реакцию циклизации для высвобождения циклического пептида (рис. 1).
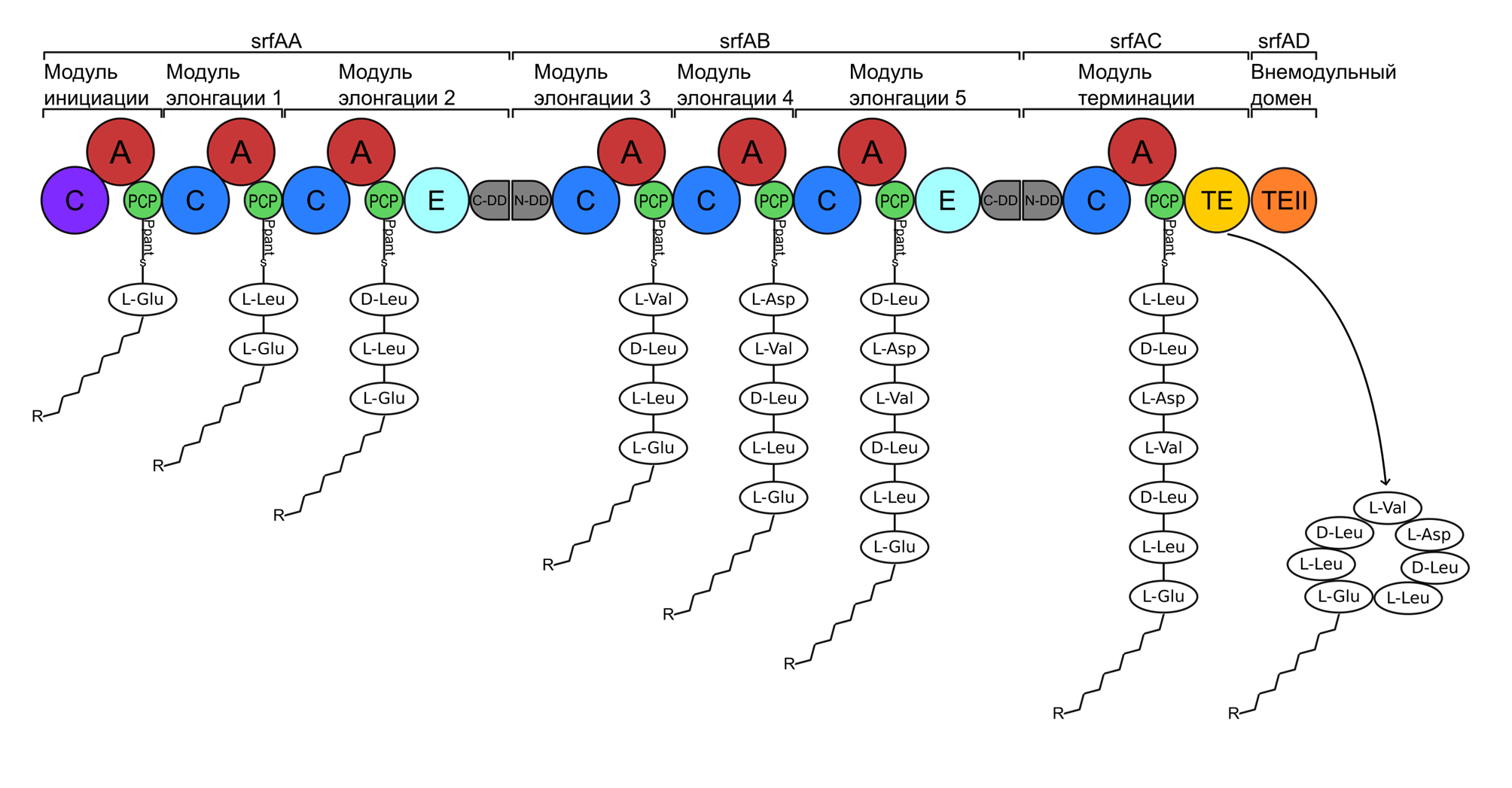
Рисунок 1. Схема нерибосомального синтеза на примере синтеза сурфактина. Домены аденилирования обозначены красными кругами, домены конденсации - синими, домены белка-переносчика пептидилов — зелеными, тиоэстеразный домен обозначен желтым кругом. Сурфактинсинтетаза состоит из трех субъединиц SrfAA, SrfAB и SrfAC, которые вместе образуют семь модулей, каждый из которых отвечает за включение одного аминокислотного остатка. Стартовый домен конденсации (фиолетовый круг) отличается от остальных тем, что катализирует N-концевое ацилирование первой аминокислоты, используя жирные кислоты. Домены эпимеризации (бирюзовые круги) обеспечивают превращение L-аминокислот в D-аминокислоты. Докинг-домены (серые фигуры) обеспечивают правильную последовательность взаимодействия между субъединицами. SrfAD кодирует, необязательную для синтеза, внешнюю тиоэстеразу TEII (оранжевый круг) с корректирующей функцией.
Помимо основных доменов, модули НРПС могут содержать дополнительные домены, например оксидазные, редуктазные, эпимеризационные и метилтрансферазные, ответственные за модификацию пептида (Miller, Gulick, 2016). Также, в ряде случаев терминальный этап синтеза может осуществляться не тиоэстеразным, а редуктазным доменом, или вообще неферментативным путем. Один из примеров неферментативного высвобождения пептида обнаружен во время биосинтеза капурамицина. Поскольку TE домен в его синтетазе отсутствовал, первоначально была выдвинута гипотеза, что домен C катализирует внутримолекулярную лактамизацию, как это происходит в CT-доменах грибковых НРПС, однако, используя синтетические тиоэфиры L -лизина в качестве контролей, было продемонстрировано, что лактамизация происходит спонтанно в присутствии воды. Результаты также поднимают интригующий вопрос о том, как другие NRPS, участвующие в биосинтезе NRP, содержащих нециклизованный L-лизин, избегают лактамизации во время процесса удлинения пептидной цепи (McErlean et al., 2019).
На генетическом уровне гены, ответственные за нерибосомальный синтез организованы в биосинтетические генные кластеры (BGC), которые содержат не только гены, кодирующие модули НРПС, но и дополнительные ферменты, транспортеры а также регуляторные гены (Jian и др., 2023).
3. Домены НРПС
3.1. А-домен
Домен аденилирования представляет собой каталитический домен, осуществляющий селективное распознавание, активацию и загрузку чрезвычайно разнообразного спектра субстратов, включающего более 500 различных мономеров, таких как непротеиногенные аминокислоты в L- и D-конфигурациях, α-, β- и δ-аминокислоты, 2-аминоизомасляную кислоту (Aib), гидроксифенилглицин (Hpg), дигидроксибензоат (Dhb) и множество других структурно разнообразных соединений нерибосомного синтеза пептидов. При этом некоторые А-домены обладают сниженной специфичностью по отношению к субстратам, например, один и тот же А-домен может аденилировать как аргинин, так и лизин Такая вариативность приводит к синтезу сразу нескольких вариантов пептидов, что, вероятно, представляет собой механизм адаптации к изменяющимся условиям окружающей среды, позволяя синтезировать несколько аналогов в зависимости от доступности субстратов(Crawford и др., 2011; Stanišić, Kries, 2019; Zhang и др., 2024).
А-домен характеризуется значительными размерами, составляющими приблизительно 500-550 аминокислотных остатков, и демонстрирует сложную двухдоменную архитектуру, включающую более крупный N-концевой субдомен Acore и меньший C-концевой субдомен Asub, которые разделены широкой щелью. Субстрат-связывающий карман локализован в пределах субдомена Acore и десятью ключевыми аминокислотными остатками, локализованными между мотивами a3 и a6, которые составляют так называемую последовательность, определяющую субстратную специфичность. Два особенно значимых консервативных остатка формируют критические водородные связи с α-аминогруппой и α-карбоксильной группой субстрата: Asp235 и каталитический остаток Lys517, тогда как остальные восемь остатков взаимодействуют с боковой цепью субстрата, обеспечивая его специфическое распознавание и правильное позиционирование в активном центре (Miller, Gulick, 2016; Nemoto и др., 2025).
Каталитический механизм А-доменов реализуется через строго регулируемую последовательность конформационных изменений, включающую четыре основные стадии, определяемые каталитическим состоянием и позиционированием субдоменов Asub и PCP относительно каталитической платформы, образованной С-доменом и субдоменом Acore. На первой стадии А-домен принимает открытую конформацию, позволяющую диффузию АТФ и субстрата в активный центр, где взаимодействие с остатками связывающего кармана обеспечивает правильное позиционирование функциональных групп.
На второй стадии субдомен Asub поворачивается приблизительно на 30°, закрываясь по направлению к субдомену Acore, что приводит каталитический лизин, расположенный на гибкой петле, внутрь активного центра и подготавливает систему к осуществлению реакции аденилирования. Заряженный каталитический лизин взаимодействует одновременно с АТФ и кислотным субстратом, сближая их в пространстве и в конечном итоге стимулируя атаку карбоксилата на α-фосфат АТФ, что приводит к образованию ацил-АМФ интермедиата (активированного субстрата) и высвобождению неорганического пирофосфата.
Третья стадия включает дальнейшее движение субдомена Asub, который поворачивается приблизительно на 140° в горизонтальной плоскости, представляя теперь противоположную поверхность субдомену Acore и перемещая PCP-домен над активным центром. Кофактор фосфопантетеин может теперь проникнуть в связывающий карман и присоединить субстрат посредством тиоэтерификации, высвобождая АМФ в процессе и подготавливая субстрат для транспорта к соседним доменам.
На заключительной четвертой стадии субдомен Asub снова поворачивается приблизительно на 180° и отдаляется от субдомена Acore, согласованно с поворотом PCP-домена, что позволяет последнему преодолеть необходимое расстояние для достижения С-домена и обеспечения субстрата для реакции конденсации, в то время как А-домен возвращается в исходную открытую конформацию, готовую к началу нового цикла (Zhang и др., 2024).
3.2. PCP-домен
Для обретения каталитической активности апо-PCP-домены должны подвергаться посттрансляционной модификации специфическими ферментами, называемыми 4'-фосфопантетеинилтрансферазами (PPTases), которые катализируют перенос 4'-фосфопантетеинового кофактора, производного кофермента A, на этот выступающий остаток серина, с образованием фосфопантеиновой «руки», способной к значительным конформационным изменениям позволяет PCP-домену взаимодействовать с множественными окружающими доменами (Duban, Cociancich, Leclère, 2022; Iacovelli, Bovenberg, Driessen, 2021).
Механизм функционирования PCP-домена включает несколько критически важных этапов: после аденилирования субстрата A-доменом происходит трансэстерификация активированного аминоацил-AMФ на сульфгидрильную группу фосфопантетеина с образованием тиоэфирной связи и высвобождением AМФ. Затем PCP-домен транспортирует ковалентно связанный субстрат к активному центру домена конденсации, где происходит образование пептидной связи между донорным и акцепторным субстратами.
Несмотря на то что A-домены рассматриваются как основные «привратники» в системах нерибосомного пептидного синтеза, накапливающиеся экспериментальные данные свидетельствуют о том, что PCP-домены также могут проявлять определенную степень специфичности по отношению к своим субстратам, что добавляет дополнительный уровень контроля качества в процесс биосинтеза.
Некоторые исследования демонстрируют, что PCP-домены из модулей, содержащих эпимеризационные домены, значительно отличаются по своей первичной структуре от стандартных PCP-доменов, обычно связанных с конденсационными доменами, что, вероятно, отражает специализированную коммуникацию с модифицирующими доменами (Corpuz, Sanlley, Burkart, 2022; Iacovelli, Bovenberg, Driessen, 2021).
3.3. С-домен и его вариации
Домены C чаще всего располагаются в начале каждого модуля НРПС и катализируют образование пептидной связи между двумя субстратами, как правило, соседних модулей. Домен конденсации состоит из двух больших субдоменов, обозначаемых как N-концевой субдомен (CNTD) и C-концевой субдомен (CCTD). Эти структурные элементы образуют характерную V-образную конфигурацию. Каждый из субдоменов содержит большую центральную β-складчатую область, окруженную несколькими α-спиральными элементами. Между двумя лопастями располагается центральный туннель, в котором находится каталитический активный сайт, содержащий консервативный мотив HHxxxDG, ответственный за образование пептидной связи между субстратами с помощью нуклеофильной атаки α-аминогруппы акцепторного субстрата на карбонильный углерод донорного тиоэфира. Согласно современным представлениям, консервативная гистидиновая группа в мотиве HHxxxDG осуществляет правильное позиционирование субстрата через образование водородных связей с α-аминогруппой акцепторного субстрата, что способствует нуклеофильной атаке на донорный карбонил и замена этого остатка на аланин полностью элиминирует каталитическую способность домена (Hur, Vickery, Burkart, 2012; Patteson и др., 2022).
Интересной структурной адаптацией является модификация канонического мотива HHxxxDG в некоторых С-доменах, например, вариант HHxxxDE, который специфически адаптирован для работы с малыми акцепторными субстратами, такими, как глицин. Глутаминовая кислота в этом варианте мотива стабилизирует и ориентирует аминогруппу акцепторного субстрата для обеспечения эффективной нуклеофильной атаки на донорный тиоэфир (Izoré и др., 2021).
Домен конденсации обладает двумя различными сайтами связывания субстратов, расположенными на противоположных сторонах центрального туннеля. Акцепторный сайт предназначен для связывания аминоацил-тиоэфира, присоединенного к соответствующему PCP-домену данного модуля, в то время как донорный сайт взаимодействует с аминоацил-тиоэфирным субстратом, переданным от предыдущего модуля. Несмотря на отсутствие четко определенных карманов субстратной специфичности, С-домены тоже участвуют в процессе корректировки синтеза, предотвращающем выполнение реакции конденсации при несоответствии субстрата, связанного с PCP-доменом, при этом акцепторный сайт демонстрирует значительно более высокую субстратную специфичность (распознавание D/L-аминокислот, N-метильных остатков, оксазолин/тиазолин-акцепторов), тогда как донорный сайт более толерантен к вариациям субстрата (Chu Yuan Kee и др., 2022; Folger и др., 2024; Iacovelli, Bovenberg, Driessen, 2021; Izoré и др., 2021).
Следует отметить большое разнообразие вариантов доменов конденсации, выполняющих разнообразные функции в нерибосомальных пептид-синтетазах. LCL- и DCL-домены представляют собой две самые распространенные подгруппы С-доменов. LCL катализирует образование пептидной связи между двумя L-конфигурированными аминокислотами. DCL-домены используют D-донор и L-акцептор. Исследования мутаций в C-доменах GrsB1 и TycB1 продемонстрировали, что замена глутамата в позиции 386 на лейцин (E386L) приводила к потере специфичности к D-субстратам и приобретению активности по L-донарам, фактически превращая DCL-домен в LCL-домен, а обратная мутация в С-домене SrfAC, превращала LCL-домен в DCL-домен (Peng и др., 2024).
Некоторые C-домены (C/E-домены) обладают бифункциональной активностью, выполняя как конденсацию, так и эпимеризацию. Их архитектура представляет собой гибрид из E-домена встроенного между N- и C-субдоменами классического C-домена, с двумя независимыми активными центрами (Wang и др., 2022). Эти домены особенно характерны для биосинтеза липопептидов у бактерий родов Pseudomonas, Burkholderia и Xanthomonas.
Стартовые C-домены (Сstart) в инициирующих модулях NRPS обеспечивает присоединение нестандартных или ацильных стартерных групп (ацил-CoA, β-гидрокси кислот и др.) к первым аминокислотам пептидной цепи. Эти домены отличаются от стандартных C-доменов расширенным донорным сайтом, способным принимать ацильные доноры (Zhong и др., 2021).
Во многих модулях НРПС C-домен заменяется, родственным ему доменом гетероциклизации (Cy-домен), который сначала катализирует образование амидной связи между донорным и акцепторным субстратами, а затем внутримолекулярную циклодегидратацию с образованием оксазолинового или тиазолинового гетероцикла из боковых цепей Ser/Thr или Cys соответственно (Duerfahrt и др., 2004). Кристаллографические и мутационные исследования показали, что в циклодегидратации участвует ранее недооценённая каталитическая диада Asp–Thr.
X-домены являются эволюционно родственными C-доменам, но каталитически неактивными из-за мутаций в консервативном мотиве HHxxxDG. Они служат платформами для рекрутирования монооксигеназ цитохрома P450, ответственных за циклизацию связанного с T-доменом растущего пептида в синтезе гликопептидов, таких как ванкомицин и теикопланин (Haslinger и др., 2015).
В исследовании биосинтеза нокардицина A был найден новый специализированный C-домен, который осуществляет интеграцию β-лактамного кольца непосредственно в процессе пептидной элонгации. Этот уникальный C-домен способен преобразовать серилтетрапептид в пентапептидный продукт, содержащий интегрированное β-лактамное кольцо, через сложный механизм, включающий дегидратацию серинового остатка с последующей внутримолекулярной циклизацией (Gaudelli, Long, Townsend, 2015; Long, Townsend, 2018; Long, Townsend, 2021).
3.4. Тиоэстеразные TE и TEII домены
Оба типа доменов относятся к классу сериновых гидролаз, что определяется наличием каталитической триады (Ser-His-Asp/Glu), делающей возможным как гидролиз, так и, при необходимости, лактамизацию или макроциклизацию продукта (Patel и др., 2023). TE-домены, как правило, располагаются внутри модуля терминации и отвечают за гидролитическое высвобождение или макроциклизацию пептидной цепи, а TEII представляют собой отдельные белки, выполняющие функцию удаления ошибочно загруженных или аномальных остатков с PCP-доменов, например, ацил-групп полипептидов неправильной длины, для поддержания работоспособности модулей. В отдельных случаях обнаружены тандемы TE: первый катализирует макроциклизацию, второй — высвобождение продукта (Kohli, Takagi, Walsh, 2002; Kotowska, Pawlik, 2014).
В процессе синтеза растущий пептид, ковалентно связанный с фосфопантеиновой «рукой» PCP-домена, переносится на каталитический серин TE-домена с образованием временного ковалентного ацил-TE промежуточного комплекса (O-ацил-сериновая связь). Этот комплекс затем подвергается нуклеофильной атаке. Атака молекулой воды приводит к гидролизу и высвобождению линейного пептида. Если же атака происходит внутримолекулярным нуклеофилом, таким как α-аминогруппа N-конца, аминогруппа боковой цепи лизина или гидроксигруппа серина/треонина, то это приводит к циклизации с образованием лактамного или лактонного кольца соответственно (Patel и др., 2023; Süssmuth, Mainz, 2017).
3.5. R-домены
Редуктазные домены представляют собой альтернативный механизм высвобождения пептидного продукта в нерибосомных пептид-синтетазах, заменяя тиоэстеразные (TE) домены. В отличие от TE-доменов, которые катализируют гидролиз тиоэфирной связи, R-домены осуществляют NAD(P)H-зависимое восстановительное высвобождение, что приводит к образованию продуктов со свободным C-концом, лишённым отрицательных зарядов. Это свойство открывает больше возможностей для последующих модификаций, таких как гликозилирование. Кроме того, образующиеся альдегидные фрагменты могут непосредственно обуславливать биологическую активность конечного продукта, как в случае мощного ингибитора протеасом — феллутамида B. Помимо образования линейных спиртов или альдегидов, некоторые R-домены способны катализировать макроциклизацию с образованием структур по типу пиразинонов, как это происходит при биосинтезе ауреузимина и ностоциклопептидов (Tietze и др., 2020).
Трёхмерная структура R-домена организована в два функциональных субдомена: N-концевой кофактор-связывающий субдомен с консервативной складкой Россмана для связывания NAD(P)H и C-концевой субстрат-связывающий субдомен, формирующий гидрофобный активный центр для взаимодействия с пептидил-тиоэфирным субстратом.
Каталитический механизм R-доменов включает 2e⁻ или 4e⁻ восстановление. Первоначальный акт 2e⁻ восстановления представляет собой шаг высвобождения, в результате которого образуется альдегидный продукт, он может быть конечным, как в случае синтеза предшественника сафрамицина А, однако в большинстве случаев он подвергается второму циклу 4e⁻ восстановления с потреблением ещё одной молекулы NAD(P)H, что приводит к образованию первичного спирта.
Второй восстановительный этап служит механизмом детоксикации, предотвращая накопление в клетке реактивных альдегидов. В некоторых системах NRPS эту вторую стадию катализирует отдельный фермент альдо/кеторедуктаза (Manavalan и др., 2010).
3.6. Домены модификации субстрата
Дополнительные домены модификации в бактериальных НРПС включают функциональные модули, которые изменяют конфигурацию, окислительно-восстановительное состояние и химические группы мономеров на PCP, тем самым расширяя структурное и функциональное разнообразие продуктов. К ним относятся домены эпимеризации (E), метилирования (M), формилирования (F), окисления (Ox) и кеторедуктазы (KR).
Домен эпимеризации в нерибосомальных пептидсинтетазах катализирует превращение L-аминокислотного остатка, в соответствующий D-энантиомер, что принципиально изменяет конформационные предпочтения, устойчивость к протеолизу и взаимодействия с мишенями у конечного пептида. Эволюционно E-домены являются потомками доменов конденсации и потому обладают схожим строением, включая консервативный мотив HhxxxDG, однако акцепторный сайт у E-доменов закрыт дополнительными остатками, которые блокируют вход в реакционный центр и препятствуют связыванию вышестоящего T-домена, поэтому, в отличие от домена C, которому нужны два субстрата для образование пептидной связи, домен E использует только один субстрат для эпимеризации (Iacovelli, Bovenberg, Driessen, 2021).
Домены метилирования представляют собой компактные каталитические модули (около 45 кДа), ответственные за введение метильных групп в структуру нерибосомальных пептидов на этапе их сборки. Метилирование пептидного остова значительно повышает гидрофобность пептида, повышая его устойчивость к протеолизу и проницаемость через клеточные мембраны. В НРПС они могут встречаться как отдельными доменами, так и быть интегрированы в домен A, между двумя основными мотивами A8 и A9. Наиболее распространенным типом модификации опосредованное М доменом является N-метилирование пептидного остова. Помимо модификации остова, M-домены могут катализировать и другие, более редкие типы метилирования: N-метилирование боковой цепи, например, диаминопропионовой кислоты в биосинтезе пениламицина; O-метилирование, как в случае генерации остатка O-Me-Ser в биосинтезе куцнеридов; S-метилирование в биосинтезе тиокоралина. C-метилирование в биосинтезе иерсиниабактина, где домен M, встроенный между доменами Cy и T, катализирует метилирование по Cα-атому тиазолинового кольца (Süssmuth, Mainz, 2017).
Домены формилирования преимущественно функционируют на стартовом этапе синтеза нерибосомального пептида и располагаются в модуле инициации в порядке F–A–PCP. Каталитический цикл начинается с активации аминокислоты A-доменом и переноса её на тиол PCP-домена. Затем PCP доставляет субстрат к каталитическому центру F-домена, который использует формилтетрагидрофолат (fTHF) в качестве кофактора для переноса формильной группы на α-аминогруппу субстрата, эта модификация устраняет положительный заряд на N-конце конечного продукта, что может быть чрезвычайно важно для конечных свойств продукта. Прекрасным примером служит линейный граммицидин, аминокислотная последовательность которого содержит исключительно гидрофобные остатки, N-конец блокирован формилированием валина, а C-конец блокирован этаноламином. Эти три особенности объясняют высокую нерастворимость в воде, но очень хорошую растворимость в различных органических растворителях, поскольку грамицидин не способен принимать суммарный заряд или образовывать цвиттер-ион при любом pH. Обычно две молекулы грамицидина димеризуются, образуя либо двойную спираль (форма «поры»), либо спиральный димер (форма «канала»), внедряются в мембрану и специфично транспортируют одновалентные катионы через билипидный слой, тем самым снижая трансмембранный ионный потенциал, при этом димеризация осуществляется «голова к голове», то есть N-концами, что было бы невозможно без формилирования (Kessler и др., 2004; Schoenafinger и др., 2006).
Домены Ox, зависимые от флавинмононуклеотида (FMN), отвечают за окисление гетероциклических промежуточных продуктов. Подобно доменам M, они обычно встроены в C-концевой субдомен A домена между консервативными мотивами A8 и A9. Ox-домены катализируют стереоселективное окисление пятичленных тиазолиновых или оксазолиновых колец, образованных доменами циклизации (Cy), до ароматических тиазолов или оксазолов соответственно. Эти домены очень консервативны и имеют приблизительно 40% идентичности последовательностей друг с другом (Schneider, Shen, Walsh, 2003; Süssmuth, Mainz, 2017).
Кетоацилредуктазные домены (KR), широко распространённые в поликетидсинтазах, в редких случаях обнаруживаются и в НРПС. KR-домены в НРПС катализируют NADPH-зависимое стереоспецифическое восстановление α-кетогруппы активированного субстрата до гидроксильной, превращая α-кетокислоту в α-гидроксикислоту. Эта реакция является ключевой для биосинтеза депсипептидов — соединений, содержащих в своей цепи как амидные, так и сложноэфирные связи. Классическими примерами таких продуктов являются цереулид и валиномицин. KR-домены в НРПС могут располагаться между A и PCP доменами, а могут находиться внутри A-домена, как домены М и Ox. В этом случае A-домен сначала активирует α-кетокислоту, а затем встроенный KR-домен восстанавливает её до α-гидроксикислоты до того, как субстрат поступит на следующий этап сборки (Alonzo, Schmeing, 2020; Ehling-Schulz и др., 2006; Süssmuth, Mainz, 2017).
3.7. Некаталитические домены
К группе некаталитических доменов относятся домены, которые не осуществляют прямого химического превращения субстратов, но обеспечивают важные функции, такие как обеспечение межмодульной коммуникации и регуляции каталитической активности. К основным некаталитическим доменам относятся MbtH-подобные белки (MLP) и докинг-домены (DD), также называемые коммуникационными доменами (COM).
MbtH-подобные белки (MLP) представляют собой небольшие (~70-80 аминокислот, молекулярная масса около 10 кДа) высококонсервативные белки, которые обнаруживаются в сотнях бактериальных НРПС, но полностью отсутствуют в геномах грибов и других эукариотических организмов (Felnagle и др., 2010). Кристаллографические исследования выявили компактную глобулярную структуру, состоящую из четырех β-нитей, образующих β-шпильку, и двух α-спиралей, при этом два строго консервативных триптофановых остатка (Trp-25 и Trp-35) формируют важный гидрофобный кластер на поверхности белка (Herbst и др., 2013).
Биохимические исследования выявили, что MLP помогают в сборке и поддержании правильной структуры А доменов и существенно стимулируют их каталитическую активность, присутствие MLP может повышать эффективность аденилирования в 5-10 раз, при этом эффект варьирует в зависимости от комбинации MLP и А домена. Особенно интересным является феномен функционального кросс-взаимодействия MLP, при котором MLP из одного биосинтетического пути может стимулировать активность НРПС из совершенно другой системы. Более того, геномные исследования выявили существование 50 «орфанных» MLP из различных бактериальных таксонов, которые кодируются генами, не локализованными вблизи каких-либо НРПС-кодирующих биосинтетических генных кластеров, что предполагает их участие в более широких регуляторных сетях. Наиболее примечательным открытием стала идентификация MLP MXAN_3118 из Myxococcus xanthus DK1622, который естественно взаимодействует с НРПС по крайней мере семи различных биосинтетических путей, включая системы биосинтеза ДК-ксантена, миксохромидов, миксовиресцинов и других вторичных метаболитов (Esquilín-Lebrón и др., 2018; Zhang и др., 2010).
Докинг-домены обеспечивают специфическую и направленную сборку многобелковых НРПС через формирование линейных массивов из белковых субъединиц. Структурная классификация докинговых доменов включает N-терминальные докинг-домены (N-DD), обладающие β-слоем с характерными петлевыми регионами, которые располагаются перед конденсационными доменами и функционируют как акцепторные элементы, и C-терминальные докинг-домены (C-DD) обладающие α-спиральной архитектурой, локализованные после PCP доменов и выполняющие донорные функции. Взаимодействие DD опосредовано связыванием «β-руки» акцепторной субъединицы с C-концевой α-спиралью донорной субъединицы (Fage и др., 2021). Специфичность коммуникационных доменов определяется комбинацией электростатических и гидрофобных взаимодействий между комплементарными поверхностями донорных и акцепторных доменов. Замена специфических аминокислотных остатков приводит к изменению партнёрской специфичности и, как следствие, к изменению спектра продуктов (Hahn, Stachelhaus, 2004).
4. Типы НРПС
Нерибосомальные пептид-синтетазы можно разделить по организации каталитических доменов, на 2 группы: модульные многодоменные ферменты (тип I) и автономные (тип II), то есть синтетазы, каталитические домены которых организованы не по модульному принципу, а представлены отдельными белками (Jaremko и др., 2020; Wang и др., 2014). Другая классификация НРПС основана на алгоритме сборки пептидной цепи и включает три основных типа: линейные (тип А), итеративные (тип В) и нелинейные (тип С). Отдельно выделяются гибридные ПКС-НРПС системы, включающие в себя не только модули нерибосомального синтеза, но и модули синтеза поликетидов (Duban, Cociancich, Leclère, 2022; Iacovelli, Bovenberg, Driessen, 2021; Süssmuth, Mainz, 2017).
Считается, что модульные НРПС составляют подавляющее большинство всех нерибосомальных пептидных синтетаз, и поэтому именно их структура была описана выше как наиболее типичная. Согласно имеющимся данным, модульные системы встречаются значительно чаще автономных систем типа II. Так, недавнее исследование показало, что только 18% кластеров генов НРПС прокариот принадлежали к типу II (Wang и др., 2014).
4.1. Линейные НРПС
Линейные НРПС представляют собой классическую модель нерибосомального синтеза, где каждый модуль используется однократно в строго последовательном порядке. В линейных системах пептидная цепь удлиняется согласно принципу колинеарности: порядок модулей в составе НРПС непосредственно соответствует последовательности аминокислот в синтезируемом продукте. Линейные НРПС могут состоять из одного белка, содержащего все модули и домены, необходимые для выполнения полного процесса синтеза, либо, из нескольких белков, каждый из которых предоставляет активированные субстраты или промежуточные продукты для поэтапной сборки конечного пептидного продукта (Sundlov и др., 2012; Watzel и др., 2021). Примером НРПС, состоящей из одного белка, является упомянутая ранее синтетаза колоссина А. Этот линейный пентадекапептид целиком синтезируется одним гигантским ферментом с молекулярной массой 1,8 МДа, кодируемым одним геном (Bode и др., 2015; Süssmuth, Mainz, 2017). Однако, значительно чаще, даже меньшие пептиды синтезируются несколькими белками. Например, сурфактин синтезируется опероном srfA, состоящим из трех генов: srfAA, srfAB и srfAC. Каждый ген кодирует отдельную субъединицу НРПС с разным числом модулей, так субъединицы SrfAA и SrfAB содержат по три модуля, а субъединица SrfAC один. Эти субъединицы совместно работают над синтезом циклического липогептапептида сурфактина (Xu и др., 2023).
Важно различать линейный режим биосинтеза и конечную структуру пептида, которая может быть либо линейной, как у колоссина А, либо циклической, как у сурфактина, хотя оба они синтезированы НРПС с линейным типом синтеза.
4.2. Итеративные НРПС
Итеративные НРПС типа В характеризуются многократным использованием одних и тех же модулей для синтеза повторяющихся структурных элементов в пептидной цепи. Накопление и хранение промежуточных продуктов в итеративных НРПС осуществляется либо Т-доменом (синтетазы конгоцидина и энниатина), способном удерживать растущий пептид посредством фосфопантетеиновой руки, либо непосредственно терминальным доменом (синтетаза грамицидина С), промежуточный продукт находится на хранении до тех пор, пока пептид не достигнет длины, подходящей для высвобождения, инициируемого Te-доменом. Классическим примером итеративного биосинтеза служит грамицидин С, циклический декапептид, синтезируемый синтетазой содержащей пять модулей. Повторное использование домена-TE позволяет дважды пройти полный цикл элонгации, создавая симметричный декапептид. Итеративный способ синтеза позволяет избегать создания чрезмерно крупных и сложных мегасинтетаз, подверженных неправильному сворачиванию и протеолизу. Так, синтетаза грамицидина С имеет массу 280 кДа, тогда как линейно функционирующая система была бы в два раза больше. Таким образом, итеративность обеспечивает экономию ресурсов и сохранение точности сборки сложных макроциклических структур (Hoyer, Mahlert, Marahiel, 2007; Kleinkauf, Gevers, Lipmann, 1969).
4.3. Нелинейные НРПС
Нелинейные НРПС представляют собой наиболее сложную и разнообразную группу нерибосомальных пептидных синтетаз. Отличительной чертой этих систем является нестандартное расположение хотя бы одного основного домена, например отсутствие А-домена в модуле, или наличие отдельных доменов, не организованных в модули. В отличие от линейных и итеративных систем, нелинейные НРПС характеризуются многократным использованием отдельных доменов, а не целых модулей, обеспечивая тем самым гораздо более сложные архитектуры и разнообразие конечных пептидов. Наиболее часто в таком режиме функционируют домены аденилирования, которые активируют и поставляют аминоацил-АМФ не только для своего родственного T-домена, но и для T-доменов других модулей в составе того же или других белков. Такое взаимодействие обеспечивается различными как междоменными взаимодействиями, так и вспомогательными факторами, такими как, тиоэстеразы II типа (TEII) и специализированные транспортные белки (Fiedler и др., 2024).
Классическим примером нелинейного биосинтеза служит система синтеза маннопептимицина. В процессе сборки этого липогликопептидного антибиотика один A-домен аденилирует β-гидроксиэндурацидидин не только для собственного T-домена, но и для T-домена следующего модуля, в котором нет собственного A-домена. Такая организация доменов с топологией C-A-T-C-T приводит к дупликации остатка эндурацидидина в растущей пептидной цепи (Magarvey и др., 2006).
В недавно описанной синтетазе фимсбактина А у Acinetobacter baumannii нелинейность распространяется на многофункциональный Cy-домен, который катализирует разветвление пептидных цепей и вторичное конденсационное событие между двумя дипептидными промежуточными продуктами, образуя тетрапептидный тиоэфир с ветвлением, тем самым обеспечивая специфическую разветвленную структуру фимсбактина А (Yang и др., 2025).
Нелинейные НРПС демонстрируют замечательную гибкость и универсальность в структуре и механизме, что делает их перспективными объектами для биоинженерии и создания новых биологически активных соединений.
5. Регуляция нерибосомального синтеза
Регуляция NRPS происходит преимущественно на уровне транскрипции генов, кодирующих эти синтетазы, и включает сложные регуляторные сети, охватывающие до 12% всего генома у некоторых микроорганизмов, таких как Streptomyces (Yan и др., 2025).
У спорообразующих бактерий, таких как представители родов Bacillus и Paenibacillus, синтез НРПС тесно связан с глобальными регуляторами спорообразования, биопленкообразования, состояния компетенции и других жизненно важных процессов в бактериальной клетке.
Так, например, Spo0A является регулятором спорообразования и одновременно ключевым активатором многих генов вторичного метаболизма, включая НРПС. Этот белок напрямую связывается с промоторными областями генов НРПС и активирует их транскрипцию в стационарной фазе роста. У Paenibacillus polymyxa было показано, что Spo0A как напрямую, так и через влияние на другие гены-регуляторы активирует экспрессию генов полимиксинового кластера (Cui и др., 2023; Liu и др., 2023).
Ген AbrB функционирует как регулятор перехода к стационарной фазе роста, контролирующий множество генов, включая многие НРПС. AbrB также связывается с промоторными областями, но при этом блокирует транскрипцию. Его активность максимальна в экспоненциальной фазе роста, а при переходе в стационарную фазу уровень AbrB снижается, что приводит к активации синтеза нерибосомальных пептидов. Сам abrB при этом контролируется Spo0A, который подавляет abrB связываясь с консенсусными Spo0A-боксами в его регуляторном регионе. Такая регуляторная система обеспечивает производство нерибосомальных пептидов в наиболее подходящее время жизненного цикла бактерии — стационарной фазе, когда уровни AbrB снижаются, а активность Spo0A возрастает. Нарушения в гене abrB приводят к сверхэкспрессии генов нерибосомального синтеза в экспоненциальной фазе (Banse и др., 2008; Li и др., 2021; Wang и др., 2024).
ComP-ComA система регулирует синтез сурфактина у бацилл в зависимости от плотности популяции. ComA фосфорилируется ComP в ответ на достижение необходимой концентрации сигнального пептида ComX и связывается с ComA-боксами в промоторе srfA оперона, инициируя транскрипцию (Piazza, Tortosa, Dubnau, 1999; Roggiani, Dubnau, 1993; Zhi, Wu, Xu, 2017).
DegS-DegU система играет центральную роль в регуляции многих НРПС, особенно у грамположительных бактерий. Белок DegU является регулятором ответа, который активируется фосфорилированием сенсорной киназой DegS. Различные уровни фосфорилирования DegU определяют его специфичность: низкие уровни фосфорилирования активируют гены подвижности и компетентности, а высокие уровни инициируют транскрипцию генов внеклеточных протеаз и НРПС. При этом фосфорилирование самого DegU регулируется несколькими различными факторами транскрипции: репрессор SinR ингибирует degU, связываясь с его промоторной областью и предотвращая инициацию транскрипции, антирепрессорный белок SlrR действует в качестве дерепрессора, способного связываться с SinR и нейтрализовать его репрессорную активность; (Ogura, Yoshikawa, Chibazakura, 2014). При этом белок DegQ функционирует как активатор фосфорилирования DegU, стимулируя активность киназы DegS и способствуя накоплению фосфорилированной формы DegU в клетке (Lilge и др., 2021; Miras, Dubnau, 2016).
При избытке азота глутаминсинтетаза GlnA находится в ингибированной форме и связываясь с TnrA подавляя его активность, при дефиците азота GlnA переходит в активную форму, активируя TnrA который, в свою очередь, связывает с промоторной областью degU и индуцирует его транскрипцию (Yasumura, Abe, Tanaka, 2008).
CcpA, глобальный регулятор транскрипции, который действует как репрессор или реже как активатор большого количества генов в зависимости от наличия предпочтительных источников углерода, в присутствии глюкозы он связывается с CRE-сайтом в промоторной области гена degU и активирует его транскрипцию (Ishii, Tanaka, Ogura, 2013).
Также, важную роль играют ион-зависимые механизмы, которые регулируют транскрипцию нерибосомально синтезируемых сидерофоров. Ключевыми для регуляции являются железозависимые репрессоры Fur/DtxR/IdeR, медь-чувствительные репрессоры CsoR/RicR а также цинк-регуляторы Zur/SmtB, опосредованно влияющие на металлотропные метаболиты (Chen и др., 2019; Chillappagari и др., 2010; Kallifidas и др., 2010; Lyu и др., 2022; Schmitt и др., 1995; Troxell, Hassan, 2013).
6. Основные подходы к инженерии НРПС
Сайт-специфический мутагенез аденилирующих доменов - наиболее стабильная, точная и хорошо изученная методика генетической инженерии НРПС. Она основана на модификации десяти ключевых аминокислотных остатков, которые определяют субстратную специфичность и расположены в активном центре A-домена, что позволяет с высокой точностью перепрограммировать распознавание аминокислот. Так, исследователи успешно модифицировали A домен модуля 10 в CdaPS3, введя единичную мутацию K278Q, которая изменила субстратное предпочтение с глутаминовой кислоты на глутамин. В другом случае единичная мутация W239S в A-домене GrsA грамицидин S синтетазы обеспечила смену специфичности с фенилаланина на O-пропаргил-тирозин (Williams, 2013; Winn и др., 2016). Третий A-домен FusA, в норме неспецифичный, принимает множественные аминокислоты (Tyr, Val, Ile, Phe), однако сайт-направленный мутагенез (S239W, L299I, G322A, V330I) перенаправил специфичность к фенилаланину, увеличив продукцию наиболее биоактивного варианта фузарицидина в три раза (Zhang и др., 2024).
Замещение доменов и модулей представляет собой более радикальный подход, включающий обмен целых доменов или модулей между различными НРПС для изменения продукта. Так, в масштабных исследованиях даптомицина компанией Cubist Pharmaceuticals с помощью обмена целых субъединиц НРПС между биосинтетическими путами даптомицина, A54145 и кальций-зависимых антибиотиков (CAD) были получены более 120 аналогов даптомицина с сохраненной или улучшенной антибактериальной активностью (Winn и др., 2016).
Замещение субдоменов представляет более тонкий подход, включающий обмен только субдоменов A-домена, ответственных за связывание субстрата, без нарушения междоменных взаимодействий. Этот подход был опробован на синтетазах грамицидина и гормаомицина, позволив создать химерные A-домены с новой специфичностью (Kries, 2016).
Вырезание и вставка целых модулей из НРПС позволяет получить пептиды с новой структурой и принципиально иными биологическими активностями, что было наглядно продемонстрировано при инженерии сурфактинсинтетазы. Целенаправленная делеция модуля 2 из трехбелковой НРПС системы сурфактина с последующим прямым соединением модулей 1 и 3 через оптимизированные линкерные последовательности приводила к успешному образованию укороченного гексапептидного варианта сурфактина без лейцинового остатка, который сохранил при этом свои поверхностно-активные свойства (Hu, Liu, Li, 2019). При инженерии балимицинсинтетазы была продемонстрирована возможность удлинения пептидной цепи через вставку специально сконструированного модуля между существующими модулями 4 и 5 белка BspB, что привело к генерации как октапептидных, так и гептапептидных продуктов, содержащих три остатка гидроксифенилглицина вместо исходных двух (Zhang и др., 2024). Следует заметить, что радикальные модификации на уровне доменов и модулей неизбежно сопровождаются значительным снижением общей эффективности биосинтеза (Kries, 2016; Williams, 2013; Winn и др., 2016; Zhang и др., 2024).
Комбинация мутагенеза и селекции является одним из наиболее эффективных методов оптимизации работы НРПС. Яркими примерами успешного применения этой методики служат работы с энтеробактин синтетазой, где замена родного A-домена EntF на домен сирингицин-синтетазы SyrE, привела к 30-кратному снижению эффективности, но после селекции эффективность синтеза у штамма была восстановлена практически до первоначального уровня (Kries, 2016).
Инженерия вспомогательных и модифицирующих ферментов представляет собой альтернативную стратегию расширения химического разнообразия продуктов НРПС, через интеграцию дополнительных ферментативных активных элементов, способных модифицировать как предшественники, так и конечные продукты биосинтеза, не затрагивая основные каталитические домены. Интеграция галогеназ из разных путей в биосинтез эндурацидина изменила паттерны хлорирования, в результате чего образовались моно-, ди- и трихлорированные варианты, а интеграция маннозилтрансферазы из синтетазы рамопланина в штамм, продуцирующих эндурацидин, привела к появлению новых мономаннозилированных вариантов эндурацидина (Winn и др., 2016).
7. Практическое применение
Фармацевтическая промышленность активно использует бактериальные НРП в качестве антибиотиков, причем несколько основных классов этих соединений стали ключевыми средствами лечения тяжелых бактериальных инфекций. В последнее время даптомицин, продуцируемый Streptomyces roseosporus, стал критически важным антибиотиком «последней линии» для борьбы с метициллин-резистентным Staphylococcus aureus (MRSA) и другими мультирезистентными патогенами. Ванкомицин, еще один важный антибиотик на основе бактериального НРП, остается незаменимым для лечения тяжелых грамположительных инфекций, особенно устойчивых к другим антибиотикам.
Разработка новых антибиотиков на основе бактериальных НРП становится все более актуальной в условиях глобального распространения антимикробной резистентности, при этом исследования сосредоточены как на открытии новых соединений, так и на модификации существующих для преодоления механизмов устойчивости (Jiang и др., 2023; Khabthani, Rolain, Merhej, 2021; Kilianova и др., 2024; Li и др., 2018a; Li и др., 2018b).
Однако применение бактериальных нерибосомальных пептидов выходит далеко за рамки антибиотиков, играя ключевую роль в различных областях медицины. Блеомицин, продуцируемый Streptomyces verticillus, является мощным противоопухолевым агентом, используемым для лечения рака яичек, лимфомы Ходжкина и других злокачественных новообразований. Этот препарат действует за счет генерации активных форм кислорода, повреждающих ДНК, что делает его особенно эффективным против быстро делящихся раковых клеток (Karpiński, Adamczak, 2018).
Бактериальные сидерофоры нашли широкое применение в сельском хозяйстве в качестве стимуляторов роста растений и агентов биоконтроля. Эти соединения улучшают усвоение железа растениями, преобразуя менее доступное трехвалентное железо (Fe³⁺) в биодоступные формы, что значительно повышает урожайность и качество культур. Штаммы Pseudomonas, продуцирующие сидерофор пиовердин, продемонстрировали высокую эффективность в стимуляции роста сахарного тростника, райграса и огурцов. Аналогичные эффекты были отмечены у Streptomyces, выделенных из рисовых растений, которые увеличивали биомассу корней и побегов (Musialowski и др., 2023; Sharma и др., 2019; Styczynski и др., 2022; Timofeeva, Galyamova, Sedykh, 2022). Сидерофор-продуцирующие бактерии, такие как Pseudomonas putida и Bacillus subtilis, эффективно подавляют вредные микроорганизмы, включая Fusarium oxysporum и Sclerotium rolfsii, снижая потребность в синтетических пестицидах (Nerek и др., 2022; Abo-Zaid и др., 2023; Moon, Sang, 2024; Zou и др., 2023).
Комбинированное применение различных штаммов B. subtilis, продуцирующих разные типы липопептидов, создает аддитивный эффект в защите растений. Например, совместное использование штаммов, продуцирующих сурфактин и итурин, значительно повышает устойчивость пшеницы к Schizaphis graminum через активацию салицилат- и этилен-зависимых защитных генов (Rumyantsev и др., 2023), а штамм B. subtilis QST 713 продемонстрировал способность ингибировать рост микотоксин-продуцирующих грибов, таких как Aspergillus flavus, Fusarium graminearum и Alternaria alternata, снижая радиальный рост до 86% и полностью подавляя продукцию афлатоксина B1, дезоксиниваленола и зеараленона (Bertuzzi и др., 2022).
8. Заключение
Нерибосомальные пептидсинтетазы являются одними из наиболее сложных и универсальных биосинтетических систем в природе, способными производить структурно разнообразные соединения с выраженной биологической активностью. Детальное понимание молекулярных механизмов функционирования отдельных доменов НРПС, от базовых каталитических до специализированных модифицирующих, создает прочную теоретическую основу для рационального дизайна и инженерии этих ферментативных комплексов.
Последние исследования архитектуры НРПС показывают, что отклонения от принципа строгой колинеарности, такие как повторное использование модулей и перераспределение функций между доменами, являются не исключением, а важным источником структурного разнообразия НРП. Итеративные системы позволяют уменьшить размер пептидсинтетаз, поддерживая высокую точность сборки повторяющихся мотивов, тогда как нелинейные архитектуры реализуют более сложную «логическую» программу сборки, в том числе за счёт переноса активированных мономеров между модулями и включения нестандартных стартеров. На этом фоне становится очевидно, что перспективность той или иной НРПС для биоинженерии определяется не только набором доменов, но и характером междоменных и межмодульных коммуникаций, включая COM-интерфейсы и другие вспомогательные ферменты. Также, регуляторные сети, контролирующие экспрессию НРПС у бактерий жёстко связаны с общим физиологическим состоянием клетки. Для инженерии продуцентов это означает, что модификация самих НРПС-доменов должна рассматриваться совместно с перенастройкой регуляторных узлов, отвечающих за инициацию и интенсивность транскрипции соответствующих кластеров.
Можно заключить, что определяющим фактором прогресса в инженерии НРПС будет являться не столько накопление отдельных примеров успешной замены доменов или модулей, сколько развитием предсказательных моделей, учитывающих специфичность A-доменов, требования к интерфейсам PCP и C доменов, роль вспомогательных белков и регуляторный контекст кластера. На стыке структурной биологии, вычислительного моделирования и направленной эволюции уже формируются подходы, которые позволяют сочетать точечный мутагенез, модульные перестройки и интеграцию вспомогательных ферментов для получения библиотек новых НРП с заданными свойствами, что особенно актуально в связи с ростом антибиотикорезистентности и спросом на экологически безопасные биопрепараты для медицины и сельского хозяйства.
Благодарности и финансирование
Публикация выполнена при поддержке программы стратегического академического лидерства «Приоритет 2030» № СП-12-23-04.
Список литературы
- Nerek E. и др. Pseudomonas spp. in biological plant protection and growth promotion // AIMSES. 2022. Т. 9. № 4. С. 493–504.
- Abo-Zaid G. A. и др. Evaluation of Bio-Friendly Formulations from Siderophore-Producing Fluorescent Pseudomonas as Biocontrol Agents for the Management of Soil-Borne Fungi, Fusarium oxysporum and Rhizoctonia solani // Agriculture. 2023. Т. 13. № 7. С. 1418.
- Agrawal S. и др. Nonribosomal Peptides from Marine Microbes and Their Antimicrobial and Anticancer Potential // Front Pharmacol. 2017. Т. 8. С. 828.
- Alonzo D. A., Schmeing T. M. Biosynthesis of depsipeptides, or Depsi: The peptides with varied generations // Protein Sci. 2020. Т. 29. № 12. С. 2316–2347.
- Baldim J. L. и др. A strategy for the identification of patterns in the biosynthesis of nonribosomal peptides by Betaproteobacteria species // Sci Rep. 2017. Т. 7. С. 10400.
- Banse A. V. и др. Parallel pathways of repression and antirepression governing the transition to stationary phase in Bacillus subtilis // Proc Natl Acad Sci U S A. 2008. Т. 105. № 40. С. 15547–15552.
- Bertuzzi T. и др. Reduction of Mycotoxigenic Fungi Growth and Their Mycotoxin Production by Bacillus subtilis QST 713 // Toxins (Basel). 2022. Т. 14. № 11. С. 797.
- Bode H. B. и др. Structure Elucidation and Activity of Kolossin A, the D-/L-Pentadecapeptide Product of a Giant Nonribosomal Peptide Synthetase // Angew Chem Int Ed Engl. 2015. Т. 54. № 35. С. 10352–10355.
- Borsetto C. и др. Microbial community drivers of PK/NRP gene diversity in selected global soils // Microbiome. 2019. Т. 7. № 1. С. 78.
- Cai X., Zhao L., Bode H. B. Engineering of Specific Single-Module Nonribosomal Peptide Synthetases of the RXP Type for the Production of Defined Peptides // ACS Synth. Biol. 2023. Т. 12. № 1. С. 203–212.
- Chen H. и др. Negative regulation of bleomycins biosynthesis by ArsR/SmtB family repressor BlmR in Streptomyces verticillus // Appl Microbiol Biotechnol. 2019. Т. 103. № 16. С. 6629–6644.
- Chillappagari S. и др. Copper Stress Affects Iron Homeostasis by Destabilizing Iron-Sulfur Cluster Formation in Bacillus subtilis // Journal of Bacteriology. 2010. Т. 192. № 10. С. 2512–2524.
- Chu Yuan Kee M.-J. и др. Structural insights into the substrate-bound condensation domains of non-ribosomal peptide synthetase AmbB // Sci Rep. 2022. Т. 12. № 1. С. 5353.
- Corpuz J. C., Sanlley J. O., Burkart M. D. Protein-protein interface analysis of the non-ribosomal peptide synthetase peptidyl carrier protein and enzymatic domains // Synth Syst Biotechnol. 2022. Т. 7. № 2. С. 677–688.
- Crawford J. M. и др. NRPS Substrate Promiscuity Diversifies the Xenematides // Org. Lett. 2011. Т. 13. № 19. С. 5144–5147.
- Cui Y. и др. Abh, AbrB3, and Spo0A play distinct regulatory roles during polymyxin synthesis in Paenibacillus polymyxa SC2 // Microbiology Spectrum. 2023. Т. 12. № 1. С. e02293-23.
- Duban M., Cociancich S., Leclère V. Nonribosomal Peptide Synthesis Definitely Working Out of the Rules // Microorganisms. 2022. Т. 10. № 3. С. 577.
- Duerfahrt T. и др. Rational Design of a Bimodular Model System for the Investigation of Heterocyclization in Nonribosomal Peptide Biosynthesis // Chemistry & Biology. 2004. Т. 11. № 2. С. 261–271.
- Ehling-Schulz M. и др. Cereulide synthetase gene cluster from emetic Bacillus cereus: Structure and location on a mega virulence plasmid related to Bacillus anthracis toxin plasmid pXO1 // BMC Microbiol. 2006. Т. 6. С. 20.
- Esquilín-Lebrón K. J. и др. An Orphan MbtH-Like Protein Interacts with Multiple Nonribosomal Peptide Synthetases in Myxococcus xanthus DK1622 // Journal of Bacteriology. 2018. Т. 200. № 21. С. 10.1128/jb.00346-18.
- Fage C. D. и др. Communication Breakdown: Dissecting the COM Interfaces between the Subunits of Nonribosomal Peptide Synthetases // ACS Catal. 2021. Т. 11. № 17. С. 10802–10813.
- Felnagle E. A. и др. Nonribosomal Peptide Synthetases Involved in the Production of Medically Relevant Natural Products // Mol. Pharmaceutics. 2008. Т. 5. № 2. С. 191–211.
- Felnagle E. A. и др. MbtH-Like Proteins as Integral Components of Bacterial Nonribosomal Peptide Synthetases // Biochemistry. 2010. Т. 49. № 41. С. 8815–8817.
- Fiedler J. и др. Direct α-Hydroxy Acid Loading onto a Bacterial Thiotemplate Assembly Line via a Multienzyme Gateway // Angew Chem Int Ed Engl. 2024. Т. 63. № 30. С. e202405165.
- Folger I. B. и др. High-throughput reprogramming of an NRPS condensation domain // Nat Chem Biol. 2024. Т. 20. № 6. С. 761–769.
- Gaudelli N. M., Long D. H., Townsend C. A. β-Lactam formation by a non-ribosomal peptide synthetase during antibiotic biosynthesis // Nature. 2015. Т. 520. № 7547. С. 383–387.
- Hahn M., Stachelhaus T. Selective interaction between nonribosomal peptide synthetases is facilitated by short communication-mediating domains // Proc Natl Acad Sci U S A. 2004. Т. 101. № 44. С. 15585–15590.
- Hartnell D. M. и др. Cyanobacterial Abundance and Microcystin Profiles in Two Southern British Lakes: The Importance of Abiotic and Biotic Interactions // Toxins (Basel). 2020. Т. 12. № 8. С. 503.
- Haslinger K. и др. X-domain of peptide synthetases recruits oxygenases crucial for glycopeptide biosynthesis // Nature. 2015. Т. 521. № 7550. С. 105–109.
- Herbst D. A. и др. Structural Basis of the Interaction of MbtH-like Proteins, Putative Regulators of Nonribosomal Peptide Biosynthesis, with Adenylating Enzymes * // Journal of Biological Chemistry. 2013. Т. 288. № 3. С. 1991–2003.
- Hoyer K. M., Mahlert C., Marahiel M. A. The Iterative Gramicidin S Thioesterase Catalyzes Peptide Ligation and Cyclization // Chem Biol. 2007. Т. 14. № 1. С. 13–22.
- Hu F., Liu Y., Li S. Rational strain improvement for surfactin production: enhancing the yield and generating novel structures // Microb Cell Fact. 2019. Т. 18. С. 42.
- Hur G. H., Vickery C. R., Burkart M. D. Explorations of Catalytic Domains in Non-Ribosomal Peptide Synthetase Enzymology // Nat Prod Rep. 2012. Т. 29. № 10. С. 1074–1098.
- Iacovelli R., Bovenberg R. A. L., Driessen A. J. M. Nonribosomal peptide synthetases and their biotechnological potential in Penicillium rubens // J Ind Microbiol Biotechnol. 2021. Т. 48. № 9–10. С. kuab045.
- Ishii H., Tanaka T., Ogura M. The Bacillus subtilis Response Regulator Gene degU Is Positively Regulated by CcpA and by Catabolite-Repressed Synthesis of ClpC // J Bacteriol. 2013. Т. 195. № 2. С. 193–201.
- Izoré T. и др. Structures of a non-ribosomal peptide synthetase condensation domain suggest the basis of substrate selectivity // Nat Commun. 2021. Т. 12. № 1. С. 2511.
- Jaremko M. J. и др. Type II Non-ribosomal Peptide Synthetase Proteins: Structure, Mechanism, and Protein-Protein Interactions // Nat Prod Rep. 2020. Т. 37. № 3. С. 355–379.
- Jian B.-S. и др. Bioinformatic Analysis Reveals both Oversampled and Underexplored Biosynthetic Diversity in Nonribosomal Peptides // ACS Chem Biol. 2023. Т. 18. № 3. С. 476–483.
- Jiang K. и др. Nonribosomal antibacterial peptides isolated from Streptomyces agglomeratus 5-1-3 in the Qinghai-Tibet Plateau // Microbial Cell Factories. 2023. Т. 22. № 1. С. 5.
- Kallifidas D. и др. The Zinc-Responsive Regulator Zur Controls Expression of the Coelibactin Gene Cluster in Streptomyces coelicolor // J Bacteriol. 2010. Т. 192. № 2. С. 608–611.
- Kalyvas J. T. и др. Broad-Spectrum Gramicidin S Derivatives with Potent Activity Against Multidrug-Resistant Gram-Negative ESKAPE Pathogens // Antibiotics. 2025. Т. 14. № 5. С. 423.
- Karpiński T. M., Adamczak A. Anticancer Activity of Bacterial Proteins and Peptides // Pharmaceutics. 2018. Т. 10. № 2. С. 54.
- Kaspar F., Neubauer P., Gimpel M. Bioactive Secondary Metabolites from Bacillus subtilis: A Comprehensive Review // J. Nat. Prod. 2019. Т. 82. № 7. С. 2038–2053.
- Kessler N. и др. The Linear Pentadecapeptide Gramicidin Is Assembled by Four Multimodular Nonribosomal Peptide Synthetases That Comprise 16 Modules with 56 Catalytic Domains * // Journal of Biological Chemistry. 2004. Т. 279. № 9. С. 7413–7419.
- Khabthani S., Rolain J.-M., Merhej V. In Silico/In Vitro Strategies Leading to the Discovery of New Nonribosomal Peptide and Polyketide Antibiotics Active against Human Pathogens // Microorganisms. 2021. Т. 9. № 11. С. 2297.
- Kilianova Z. и др. Recent Trends in Therapeutic Drug Monitoring of Peptide Antibiotics // J Sep Sci. 2024. Т. 47. № 19. С. e202400583.
- Kleinkauf H., Gevers W., Lipmann F. INTERRELATION BETWEEN ACTIVATION AND POLYMERIZATION IN GRAMICIDIN S BIOSYNTHESIS* // Proc Natl Acad Sci U S A. 1969. Т. 62. № 1. С. 226–233.
- Kohli R. M., Takagi J., Walsh C. T. The thioesterase domain from a nonribosomal peptide synthetase as a cyclization catalyst for integrin binding peptides // Proc Natl Acad Sci U S A. 2002. Т. 99. № 3. С. 1247–1252.
- Komaki H. и др. Genome-based analysis of non-ribosomal peptide synthetase and type-I polyketide synthase gene clusters in all type strains of the genus Herbidospora // BMC Research Notes. 2015. Т. 8. № 1. С. 548.
- Kooner A., Soby S. Draft Genome Sequence of Pseudomonas sp. Strain MWU13-3659, Isolated from Commercial Cranberry Bog Soil in Massachusetts, USA // Microbiology Resource Announcements. 2022. Т. 11. № 11. С. e00880-22.
- Kotowska M., Pawlik K. Roles of type II thioesterases and their application for secondary metabolite yield improvement // Appl Microbiol Biotechnol. 2014. Т. 98. № 18. С. 7735–7746.
- Krätzschmar J., Krause M., Marahiel M. A. Gramicidin S biosynthesis operon containing the structural genes grsA and grsB has an open reading frame encoding a protein homologous to fatty acid thioesterases // Journal of Bacteriology. 1989. Т. 171. № 10. С. 5422–5429.
- Kries H. Biosynthetic engineering of nonribosomal peptide synthetases // J Pept Sci. 2016. Т. 22. № 9. С. 564–570.
- Léger G. и др. In Tuber Biocontrol of Potato Late Blight by a Collection of Phenazine-1-Carboxylic Acid-Producing Pseudomonas spp. // Microorganisms. 2021. Т. 9. № 12. С. 2525.
- Li Y. и др. Fusaricidin Biosynthesis Is Controlled via a KinB-Spo0A-AbrB Signal Pathway in Paenibacillus polymyxa WLY78 // MPMI. 2021. Т. 34. № 12. С. 1378–1389.
- Li Y.-X. и др. Resistance to nonribosomal peptide antibiotics mediated by d-stereospecific peptidases // Nat Chem Biol. 2018a. Т. 14. № 4. С. 381–387.
- Li Y.-X. и др. Discovery of cationic nonribosomal peptides as Gram-negative antibiotics through global genome mining // Nat Commun. 2018b. Т. 9. С. 3273.
- Lilge L. и др. Expression of degQ gene and its effect on lipopeptide production as well as formation of secretory proteases in Bacillus subtilis strains // Microbiologyopen. 2021. Т. 10. № 5. С. e1241.
- Liu N. и др. Transcription Factor Spo0A Regulates the Biosynthesis of Difficidin in Bacillus amyloliquefaciens // Microbiology Spectrum. 2023. Т. 11. № 4. С. e01044-23.
- Long D. H., Townsend C. A. Mechanism of Integrated β‑Lactam Formation by a Nonribosomal Peptide Synthetase during Antibiotic Synthesis // Biochemistry. 2018. Т. 57. № 24. С. 3353–3358.
- Long D. H., Townsend C. A. Acyl Donor Stringency and Dehydroaminoacyl Intermediates in β-Lactam Formation by a Non-ribosomal Peptide Synthetase // ACS Chem. Biol. 2021. Т. 16. № 5. С. 806–812.
- Lyu M. и др. Zinc-Responsive Regulator Zur Regulates Zinc Homeostasis, Secondary Metabolism, and Morphological Differentiation in Streptomyces avermitilis // Applied and Environmental Microbiology. 2022. Т. 88. № 7. С. e00278-22.
- Mach B., Reich E., Tatum E. L. Separation of the biosynthesis of the antibiotic polypeptide tyrocidine from protein biosynthesis* // Proceedings of the National Academy of Sciences. 1963. Т. 50. № 1. С. 175–181.
- Magarvey N. A. и др. Biosynthetic Pathway for Mannopeptimycins, Lipoglycopeptide Antibiotics Active against Drug-Resistant Gram-Positive Pathogens // Antimicrob Agents Chemother. 2006. Т. 50. № 6. С. 2167–2177.
- Manavalan B. и др. Molecular modeling of the reductase domain to elucidate the reaction mechanism of reduction of peptidyl thioester into its corresponding alcohol in non-ribosomal peptide synthetases // BMC Struct Biol. 2010. Т. 10. С. 1.
- Mansour B., Gauld J. W. Computational Insights into Amide Bond Formation Catalyzed by the Condensation Domain of Nonribosomal Peptide Synthetases // ACS Omega. 2024. Т. 9. № 26. С. 28556–28563.
- McErlean M., Overbay J., Van Lanen S. Refining and expanding nonribosomal peptide synthetase function and mechanism // J Ind Microbiol Biotechnol. 2019. Т. 46. № 3–4. С. 493–513.
- Miller B. R., Gulick A. M. Structural Biology of Non-Ribosomal Peptide Synthetases // Methods Mol Biol. 2016. Т. 1401. С. 3–29.
- Miras M., Dubnau D. A DegU-P and DegQ-Dependent Regulatory Pathway for the K-state in Bacillus subtilis // Front. Microbiol. 2016. Т. 7.
- Moon H. J., Sang M. K. Biocontrol of Southern Blight Caused by Sclerotium rolfsii in Pepper Plants Using Bacillus subtilis GJ6-14 // Res. Plant Dis. 2024. Т. 30. № 2. С. 181–188.
- Musialowski M. и др. Metabolically versatile psychrotolerant Antarctic bacterium Pseudomonas sp. ANT_H12B is an efficient producer of siderophores and accompanying metabolites (SAM) useful for agricultural purposes // Microb Cell Fact. 2023. Т. 22. С. 85.
- Nemoto M. и др. Identification of Key Amino Acids in the A Domains of Polymyxin Synthetase Responsible for 2,4-Diaminobutyric Acid Adenylation in Paenibacillus polymyxa NBRC3020 Strain // ACS Chem. Biol. 2025. Т. 20. № 2. С. 321–331.
- Ogura M., Yoshikawa H., Chibazakura T. Regulation of the Response Regulator Gene degU through the Binding of SinR/SlrR and Exclusion of SinR/SlrR by DegU in Bacillus subtilis // J Bacteriol. 2014. Т. 196. № 4. С. 873–881.
- Patel K. D. и др. Structural advances toward understanding the catalytic activity and conformational dynamics of modular nonribosomal peptide synthetases // Nat Prod Rep. 2023. Т. 40. № 9. С. 1550–1582.
- Patteson J. B. и др. Structure and Function of a Dehydrating Condensation Domain in Nonribosomal Peptide Biosynthesis // J Am Chem Soc. 2022. Т. 144. № 31. С. 14057–14070.
- Peng H. и др. Controlling Substrate- and Stereospecificity of Condensation Domains in Nonribosomal Peptide Synthetases // ACS Chem. Biol. 2024. Т. 19. № 3. С. 599–606.
- Piazza F., Tortosa P., Dubnau D. Mutational Analysis and Membrane Topology of ComP, a Quorum-Sensing Histidine Kinase of Bacillus subtilis Controlling Competence Development // J Bacteriol. 1999. Т. 181. № 15. С. 4540–4548.
- Ranjan A. и др. Nature’s Antimicrobial Arsenal: Non-Ribosomal Peptides from PGPB for Plant Pathogen Biocontrol // Fermentation. 2023. Т. 9. № 7. С. 597.
- Roggiani M., Dubnau D. ComA, a phosphorylated response regulator protein of Bacillus subtilis, binds to the promoter region of srfA. // J Bacteriol. 1993. Т. 175. № 10. С. 3182–3187.
- Rumyantsev S. D. и др. Additive Effect of the Composition of Endophytic Bacteria Bacillus subtilis on Systemic Resistance of Wheat against Greenbug Aphid Schizaphis graminum Due to Lipopeptides // Life. 2023. Т. 13. № 1. С. 214.
- Saiyam D. и др. Lipopeptides from Bacillus: unveiling biotechnological prospects—sources, properties, and diverse applications // Braz J Microbiol. 2024. Т. 55. № 1. С. 281–295.
- Schmitt M. P. и др. Characterization of an iron-dependent regulatory protein (IdeR) of Mycobacterium tuberculosis as a functional homolog of the diphtheria toxin repressor (DtxR) from Corynebacterium diphtheriae // Infection and Immunity. 1995. Т. 63. № 11. С. 4284–4289.
- Schneider T. L., Shen B., Walsh C. T. Oxidase Domains in Epothilone and Bleomycin Biosynthesis: Thiazoline to Thiazole Oxidation during Chain Elongation // Biochemistry. 2003. Т. 42. № 32. С. 9722–9730.
- Schoenafinger G. и др. Formylation Domain: An Essential Modifying Enzyme for the Nonribosomal Biosynthesis of Linear Gramicidin // J. Am. Chem. Soc. 2006. Т. 128. № 23. С. 7406–7407.
- Sharma S. и др. Foliar Application of Iron Fortified Bacteriosiderophore Improves Growth and Grain Fe Concentration in Wheat and Soybean // Indian J Microbiol. 2019. Т. 59. № 3. С. 344–350.
- Sharma V., Kaur R., Salwan R. Streptomyces: host for refactoring of diverse bioactive secondary metabolites // 3 Biotech. 2021. Т. 11. № 7. С. 340.
- Shi C. и др. Design, Synthesis, and Biophysical Evaluation of Mechanism-Based Probes for Condensation Domains of Nonribosomal Peptide Synthetases // ACS Chem. Biol. 2020. Т. 15. № 7. С. 1813–1819.
- Sieber A. и др. Fundamentals of bio-based technologies for selective metal recovery from bio-leachates and liquid waste streams // Front. Bioeng. Biotechnol. 2025. Т. 12.
- Stanišić A., Kries H. Adenylation Domains in Nonribosomal Peptide Engineering // Chembiochem. 2019. Т. 20. № 11. С. 1347–1356.
- Styczynski M. и др. Application of Psychrotolerant Antarctic Bacteria and Their Metabolites as Efficient Plant Growth Promoting Agents // Front Bioeng Biotechnol. 2022. Т. 10. С. 772891.
- Sundlov J. A. и др. Structural and functional investigation of the intermolecular interaction between NRPS adenylation and carrier protein domains // Chem Biol. 2012. Т. 19. № 2. С. 188–198.
- Süssmuth R. D., Mainz A. Nonribosomal Peptide Synthesis-Principles and Prospects // Angew Chem Int Ed Engl. 2017. Т. 56. № 14. С. 3770–3821.
- Tietze A. и др. Nonribosomal Peptides Produced by Minimal and Engineered Synthetases with Terminal Reductase Domains // Chembiochem. 2020. Т. 21. № 19. С. 2750–2754.
- Timofeeva A. M., Galyamova M. R., Sedykh S. E. Bacterial Siderophores: Classification, Biosynthesis, Perspectives of Use in Agriculture // Plants (Basel). 2022. Т. 11. № 22. С. 3065.
- Troxell B., Hassan H. M. Transcriptional regulation by Ferric Uptake Regulator (Fur) in pathogenic bacteria // Front Cell Infect Microbiol. 2013. Т. 3. С. 59.
- Walsh C. T., O’Brien R. V., Khosla C. Nonproteinogenic amino acid building blocks for nonribosomal peptide and hybrid polyketide scaffolds // Angew Chem Int Ed Engl. 2013. Т. 52. № 28. С. 7098–7124.
- Wang H. и др. Atlas of nonribosomal peptide and polyketide biosynthetic pathways reveals common occurrence of nonmodular enzymes // Proc Natl Acad Sci U S A. 2014. Т. 111. № 25. С. 9259–9264.
- Wang J. и др. Catalytic trajectory of a dimeric nonribosomal peptide synthetase subunit with an inserted epimerase domain // Nat Commun. 2022. Т. 13. № 1. С. 592.
- Wang M. и др. An Integrated Pipeline and Overexpression of a Novel Efflux Transporter, YoeA, Significantly Increases Plipastatin Production in Bacillus subtilis // Foods. 2024. Т. 13. № 11. С. 1785.
- Waongo B. и др. Prospection for potential new non-ribosomal peptide gene clusters in Bacillus genus isolated from fermented foods and soil through genome mining // Front. Microbiol. 2025. Т. 16.
- Watzel J. и др. NMR resonance assignments for a docking domain pair with an attached thiolation domain from the PAX peptide-producing NRPS from Xenorhabdus cabanillasii // Biomol NMR Assign. 2021. Т. 15. № 1. С. 229–234.
- Weckermann R., Fürbass R., Marahiel M. A. Complete nucleotide sequence of the tycA gene coding the tyrocidine synthetase 1 from Bacillus brevis. // Nucleic Acids Res. 1988. Т. 16. № 24. С. 11841.
- Williams G. J. Engineering polyketide synthases and nonribosomal peptide synthetases // Current Opinion in Structural Biology. 2013. Т. 23. № 4. С. 603–612.
- Winn M. и др. Recent advances in engineering nonribosomal peptide assembly lines // Nat. Prod. Rep. 2016. Т. 33. № 2. С. 317–347.
- Xu Y. и др. Genome-Wide Identification and Evolutionary Analyses of SrfA Operon Genes in Bacillus // Genes. 2023. Т. 14. № 2. С. 422.
- Yan H. и др. A rational multi-target combination strategy for synergistic improvement of non-ribosomal peptide production // Nat Commun. 2025. Т. 16. № 1. С. 1883.
- Yang J. и др. Chemical Logic of Peptide Branching by Iterative Nonlinear Nonribosomal Peptide Synthetases // Biochemistry. 2025. Т. 64. № 3. С. 719–734.
- Yasumura A., Abe S., Tanaka T. Involvement of Nitrogen Regulation in Bacillus subtilis degU Expression // J Bacteriol. 2008. Т. 190. № 15. С. 5162–5171.
- Zhang M. и др. Functional Diversity and Engineering of the Adenylation Domains in Nonribosomal Peptide Synthetases // Mar Drugs. 2024. Т. 22. № 8. С. 349.
- Zhang W. и др. Activation of the Pacidamycin PacL Adenylation Domain by MbtH-Like Proteins // Biochemistry. 2010. Т. 49. № 46. С. 9946–9947.
- Zhi Y., Wu Q., Xu Y. Genome and transcriptome analysis of surfactin biosynthesis in Bacillus amyloliquefaciens MT45 // Sci Rep. 2017. Т. 7. С. 40976.
- Zhong L. и др. Engineering and elucidation of the lipoinitiation process in nonribosomal peptide biosynthesis // Nat Commun. 2021. Т. 12. С. 296.
- Zou L. и др. Biocontrol and plant growth promotion potential of endophytic Bacillus subtilis JY-7-2L on Aconitum carmichaelii Debx. // Front. Microbiol. 2023. Т. 13.
Статья поступила в редакцию 11 февраля 2026 г.
Поступила после доработки 26 февраля 2026 г.
Принята к печати 15 марта 2026 г.
Received 11, February, 2026
Revised 26, February, 2026
Accepted 15, March, 2026