Введение
В настоящее время особую ценность для исследователей представляет анализ некодирующих частей генома, включая промоторы, энхансеры и другие регуляторные элементы, которые контролируют экспрессию генов и определяют пространственно-временные транскрипционные программы в различных тканях, включая гонады [4]. Понимание регуляции этих участков особенно важно для репродуктивной биологии: при высоком сходстве набора генов у человека и мыши наблюдаются принципиально различающиеся репродуктивные параметры, в частности число овулирующих фолликулов за цикл [1,4]. У взрослой мыши в одном эстральном цикле обычно овулирует около 8–10 ооцитов, тогда как у человека в норме формируется один доминантный фолликул и происходит моноовуляция; множественная овуляция рассматривается как относительно редкое отклонение. Эти различия представляют собой эволюционно закреплённый, видоспецифичный компонент репродуктивной стратегии, что обосновывает сравнительный анализ геномов для выявления регуляторных основ межвидовых вариаций [4].
Фолликулогенез играет ключевую роль в формировании функционального репродуктивного потенциала, определяя овариальный резерв, цикличность овуляции и общий гормональный фон женской репродуктивной системы [1,7]. Антимюллеров гормон (АМГ) является одним из ключевых маркеров и регуляторов роста фолликулов, участвуя в ограничении активации примордиального пула и модуляции чувствительности растущих фолликулов к внешним сигналам. Нарушения в регуляции АМГ ассоциированы с изменениями овариального резерва, риском преждевременного истощения яичников и различными формами репродуктивной дисфункции [2,3,7,9].
Ген AMН занимает центральное место в регуляции ранних этапов фолликулогенеза, а его экспрессия и тканевая специфичность определяются совокупностью промоторных, энхансерных и других цис-регуляторных элементов [5,7,8,10]. Регуляторные последовательности этого гена могут содержать как эволюционно консервативные мотивы, обеспечивающие базовые функции репродуктивной системы млекопитающих, так и видоспецифичные элементы, возникшие в ходе дивергенции линий приматов и грызунов [4]. Сравнительный анализ нуклеодитных последовательностей гена AMН у человека (Homo sapiens, hg39) и мыши (Mus musculus, mm39) позволяет выявить консервативные мотивы, ответственные за основные регуляторные функции, а также вариации, объясняющие различия в числе овуляторных фолликулов.
Цель настоящего исследования — Сравнение некодирующих регуляторных элементов гена AMH у человека и мыши: консервативные и видоспецифичные кандидаты, потенциально влияющие на число овуляторных фолликулов
Материалы и методы
Геномные последовательности человека и мыши были получены из UCSC Genome Browser (hg39 и mm39). Для каждого гена были определены координаты полных локусов, включая промоторные, интронные и возможные области энхансеров, чтобы охватить все потенциально функциональные некодирующие элементы генома. С использованием командной строки bedtools getfasta были извлечены полные нуклеотидные поседовательности гена AMH для человека и мыши. Для каждого гена были сформированы отдельные FASTA-файлы, которые затем объединялись для дальнейшего анализа. Выравнивание последовательностей проводилось с помощью MAFFT, что позволило выявить позиции полной консервации и локальные вариации в пределах регуляторных и кодирующих областей. Анализ консервативности и нуклеотидного состава учитывал степень консервативности оценивалась парной процентной идентичностью между последовательностями человека и мыши. Для локального анализа использовалось скользящее окно размером 100 нуклеотидов с шагом 50 нуклеотидов. Для каждого гена рассчитывался GC-состав каждой последовательности. Подряд идущие полностью консервативные участки длиной ≥ 6 нуклеотидов рассматривались как кандидаты на функциональные регуляторные элементы. Для выявления повторяющихся триплетов (3-mer) и потенциальных регуляторных мотивов проводился подсчёт частот всех 3-мерных последовательностей в каждой из изучаемых последовательностей (K-mer анализ). Все вычислительные процедуры и визуализация выполнялись с использованием Python3 и библиотек Biopython, Pandas, NumPy и Matplotlib.
Результаты исследований.
В таблице 1 представлены результаты выравнивания гена AMH у человека (chr19) и мыши (chr10). Нуклеотидная последовательность гена AMH (человек: chr19:2249323-2252073, длина 2751 bp; мышь: chr10:80641074-80643513, длина 2439 bp) показал общую длину выравнивания 2769 bp с 2421 сравнимой позицией, 1641 совпадением и 67.782% парной идентичности.
Таблица 1 Результаты выравнивании гена AMH у человека (chr19) и мыши (chr10)
|
Ген |
Локусы |
Длина выравнивания |
Совпало |
Число позиций выравнивания |
Пропуски |
Идентичность |
|
AMH |
chr19:2249323-2252073 |
2769 |
1641 |
2421 |
348 |
67.782 |
|
chr10:80641074-80643513 |
Сравнительный анализ нуклеотидного состава показал выраженные различия между последовательностями AMH у человека и мыши (рис. 1). При исключении позиций с пропусками выявлено, что человеческая последовательность характеризуется заметно более высоким содержанием нуклеотидов C и G. Такая повышенная GC-насыщенность обычно связана с увеличенной стабильностью двуцепочечной структуры ДНК, а также может коррелировать с особенностями транскрипционной активности и плотностью CpG-островков. Для функционально значимых генов, включая гормональные, высокая доля C/G нередко служит признаком поддерживающего отбора, направленного на сохранение консервативных регуляторных и структурных участков.
В противоположность этому, в мышиной последовательности доминируют нуклеотиды A и T, что приводит к более низкой общей GC-составляющей. Подобный сдвиг в сторону AT-богатых участков может отражать как видоспецифические особенности геномной архитектуры грызунов, так и различия в локальной мутационной среде. Известно, что геномы мышей в среднем более AT-насыщены по сравнению с человеческим геномом, что согласуется с выявленным распределением оснований в анализируемом фрагменте AMH. Более высокое содержание A и T также может быть следствием иной динамики репарации и репликации ДНК или различий в структуре некодирующих участков, соседствующих с геном.
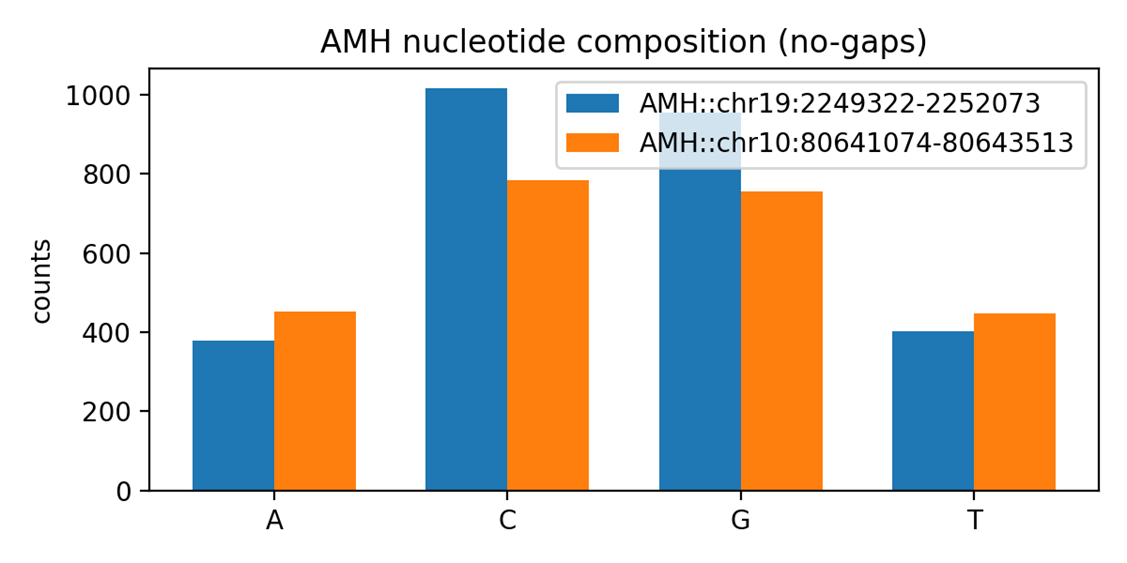
Рисунок 1. Нуклеотидный состав гена AMH
Кодирующие и ключевые регуляторные области демонстрируют высокую консервацию, в то время как интронные сегменты содержат значительное число вставок/делеций (348 пропусков). Выявлено 77 полностью консервативных участков длиной ≥ 6 bp (общая длина 767 bp), с преобладанием GC-богатых последовательностей в 5'- и 3'-областях.
Таблица 2.
Мотивы с наибольшей консервативностью ( ≥ 15 нуклеотидов) в генах АМН мыши и человека
|
Начало |
Конец |
Длина |
Последовательность |
|
2590 |
2628 |
38 |
GAGGAGCGCATCAGCGCGCACCACGTGCCCAACATGGT |
|
2557 |
2589 |
32 |
GCCTACGCGGGCAAGCTGCTCATCAGCCTGTC |
|
2171 |
2193 |
22 |
CCCCGCTGCTGGCGCGCCTGCT |
|
2482 |
2502 |
20 |
AACCACGTGGTGCTGCTGCT |
|
294 |
312 |
18 |
AGACCTGGCCACCTTCGG |
|
1655 |
1673 |
18 |
CACGGCCAGCTGGACACC |
|
2353 |
2370 |
17 |
TGCGCGCTGCGCGAGCT |
|
2539 |
2556 |
17 |
CCCTGCTGCGTGCCCAC |
|
2145 |
2160 |
15 |
CCTCCCGGGTCTGCC |
|
2254 |
2269 |
15 |
AAGGCGCTGCAGGGC |
Наиболее длинный мотив включает 38-нуклеотидную последовательность GAGGAGCGCATCAGCGCGCACCACGTGCCCAACATGGT (poz. 2590-2628). Короткие мотивы (6-12 bp) кластеризуются в промоторной области (poz. 56-375), указывая на сохранённые сайты транскрипционных факторов.
Таблица 3.
Мотивы с наименьшей консервативностью гена AMH мыши и человека
|
Начало |
Конец |
Длина |
Последовательность |
|||
|
74 |
81 |
7 |
CTGAGGC |
|||
|
152 |
159 |
7 |
GCAGCCC |
|||
|
243 |
250 |
7 |
CTATGAG |
|||
|
313 |
320 |
7 |
GTCTGCA |
|||
|
418 |
425 |
7 |
GAAGGTA |
|||
|
1119 |
1126 |
7 |
AGGTCAC |
|||
|
1170 |
1177 |
7 |
GCATGGG |
|||
|
1403 |
1410 |
7 |
GCGGCTC |
|||
|
1620 |
1627 |
7 |
TGCTGCC |
|||
|
1632 |
1639 |
7 |
CCGAGCC |
|||
|
1679 |
1686 |
7 |
TTCCCGC |
|||
|
1692 |
1699 |
7 |
GGTGCGC |
|||
|
1870 |
1877 |
7 |
CTGCGGG |
|||
|
1934 |
1941 |
7 |
GCTTCCC |
|||
|
2056 |
2063 |
7 |
CTGCACG |
|||
|
2278 |
2285 |
7 |
GAGTGGC |
|||
|
2506 |
2513 |
7 |
ATGCAGG |
|||
|
2524 |
2531 |
7 |
GCCCTGG |
|||
|
141 |
147 |
6 |
CTGGCC |
|||
|
334 |
340 |
6 |
CAGGCT |
|||
|
1011 |
1017 |
6 |
CAGTGA |
|||
|
1042 |
1048 |
6 |
GTTCCA |
|||
|
1378 |
1384 |
6 |
GTGGAC |
|||
|
1506 |
1512 |
6 |
AGCCAG |
|||
|
1555 |
1561 |
6 |
GCTGCA |
|||
|
1585 |
1591 |
6 |
CCGCTG |
|||
|
1788 |
1794 |
6 |
TTCCAG |
|||
|
1824 |
1830 |
6 |
CAGCGC |
|||
|
1987 |
1993 |
6 |
CTCGAC |
|||
Сравнительный анализ структорно-функциональной организации генов показал, что наиболее консервативные участки расположены именно в кодирующих областях. Видимые различия между человеческой и мышиной некодирующими последовательностями AMH в выравнивании могут отражать видоспецифические особенности регуляции фолликулогенеза. У человека и мыши существенно различается суммарный объём овариального резерва и динамика его расходования, эти физиологические отличия нередко сопровождаются различиями в генах, участвующих в контроле рекрутирования и созревания фолликулов. Поскольку AMH напрямую влияет на скорость активации фолликулов, различия в его нуклеотидной последовательности, в том числе в уровне консервативности отдельных участков, могут отражать адаптацию гормона к разному количеству фолликулов и различной скорости их созревания у изучаемых видов.
Таблица 4. Консервативные участки гена AMH, расположенные в некодирующих областях
|
Ген |
Начало |
Конец |
Длина |
Последовательность |
|
AMH |
418 |
425 |
7 |
GAAGGTA |
|
AMH |
942 |
951 |
9 |
GGACAGATC |
|
AMH |
1154 |
1164 |
10 |
CAGGTACCAG |
|
AMH |
1170 |
1177 |
7 |
GCATGGG |
|
AMH |
1444 |
1453 |
9 |
GGTAGGTCC |
|
AMH |
1506 |
1512 |
6 |
AGCCAG |
|
AMH |
1515 |
1526 |
11 |
CGTGCCCACCC |
|
AMH |
1692 |
1699 |
7 |
GGTGCGC |
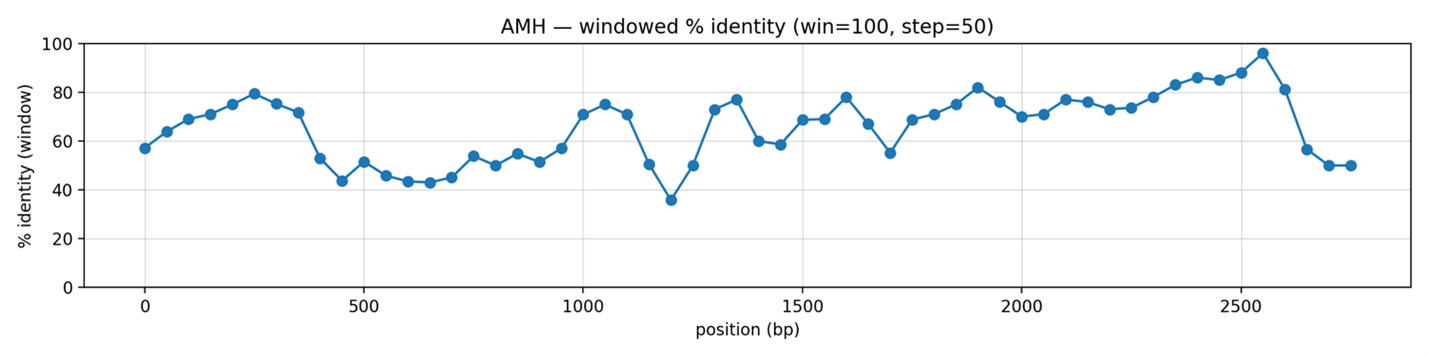
Рисунок 2. График скользящей консервации гена AMH и GC-состав
Скользящий анализ процентной идентичности (окно 100 п.н., шаг 50 п.н.) показывает, что уровень последовательного сходства между человеческим и мышиным AMH неоднороден по длине выровненного региона. В начальной части последовательности идентичность колеблется в диапазоне 55–80%, достигая локального максимума около 80% в области ~250–300 п.н. Центральная часть выравнивания характеризуется пониженной идентичностью, снижаясь до 40–45% в промежутке примерно 450–650 п.н., что указывает на наличие участков значительной дивергенции. Далее процент идентичности частично восстанавливается, достигая 70–80% в нескольких регионах, однако остаётся волнообразным и отражает чередование более консервативных и более вариабельных доменов. Вблизи конца выровненного участка наблюдается повторное снижение идентичности, что может быть связано с структурными различиями, вставками или делениями, специфичными для одного из видов.
Такая картина соответствует типичному профилю эволюционно дивергировавших генов: наличие высоко-консервативных доменов (предположительно функционально значимых) перемежается с изменчивыми участками, в которых допустима большая степень нуклеотидных замен.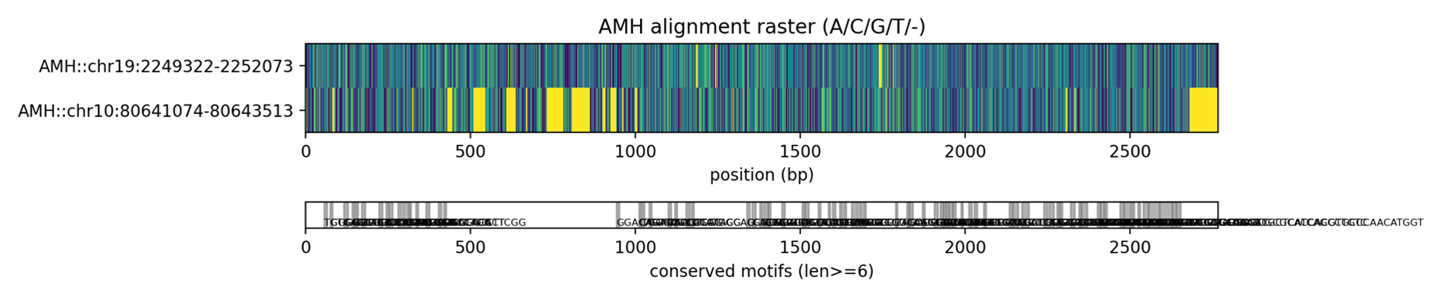
Рисунок 3. Карта мотивов гена AMH у мыши и человека.
Визуализация двух выровненных последовательностей AMH демонстрирует распределение совпадающих и различающихся нуклеотидов между человеком и мышью. Многочисленные разноцветные вертикальные колонки отражают участки расхождений, тогда как полосы более однородного окрашивания соответствуют зонам высокой консервации. В нескольких регионах, особенно в первой трети выравнивания, наблюдаются скопления вариабельных позиций, что согласуется с результатами скользящего анализа идентичности.
Под растровым изображением ( рис.3) представлена карта консервативных мотивов, выделенных по критерию минимальной длины (len ≥ 6). Эти мотивы формируют дискретные блоки, распределённые вдоль последовательности, и их локализация в значительной степени коррелирует с областями повышенной идентичности, выявленными методом скользящего окна. Отдельные мотивы располагаются как в начале, так и в середине и конце гена, что указывает на наличие нескольких функционально значимых фрагментов, сохранённых в ходе эволюции обоих видов. Полученные результаты свидетельствуют о том, что ген AMH у человека и мыши демонстрирует выраженные нуклеотидные и структурные отличия, включая вариабельные участки с пониженной идентичностью, чередующиеся с консервативными доменами. Скользящий анализ идентичности выявил несколько регионов существенной дивергенции, а растровое выравнивание и карта мотивов подтвердили наличие протяжённых участков, в которых последовательности двух видов расходятся как по составу отдельных нуклеотидов, так и по локальной структуре соответствующих элементов. Эти молекулярные различия хорошо согласуются с известными межвидовыми особенностями фолликулогенеза. У человека AMH играет ключевую роль в длительном и постепенном росте антральных фолликулов, а также в поддержании относительно продолжительного репродуктивного окна. У мыши же AMH участвует в регулировании гораздо более быстрого, циклического и динамичного фолликулогенеза, сопровождаемого более высокой скоростью рекрутирования и атрезии фолликулов. Выявленные различия в последовательности, особенно в регионах, соответствующих регуляторным или функциональным доменам гена, могут отражать адаптацию к этим разным режимам развития фолликулов.
Заключение
Таким образом, дивергентные участки гена AMH, характеризующиеся пониженной идентичностью и видоспецифическими мотивами, вероятно, способствуют формированию отличий в чувствительности фолликулов к AMH, скорости фолликулярного роста и особенностям контроля рекрутирования. Консервативные же домены обеспечивают сохранение базовой функции гормона как ингибитора ранних этапов фолликулогенеза. В совокупности эти данные указывают на то, что межвидовые вариации в последовательности AMH могут быть одним из молекулярных факторов, лежащих в основе различий в динамике и архитектуре фолликулогенеза у человека и мыши.
Финансирование. Исследование выполнено при финансовой поддержке Российского научного фонда, проект № 23–15-00464.
Список использованной литературы
- Адамян Л.В., Сибирская Е.В., Пивазян Л.Г. и др. Современный взгляд на фолликулогенез и методы его активации. Эффективная фармакотерапия. 2025; 21 (20): 84–92. DOI 10.33978/2307-3586-2025-21-20-84-92. Эффективная фармакотерапия. 2025. Том 21. № 20. Акушерство и гинекология
- Найдукова А.А., Каприна Е.К., Иванец Т.Ю., Чернуха Г.Е. Значение антимюллерова гормона в диагностике синдрома поликистозных яичников. Акушерство и гинекология. 2017; 1: 46-52.
- Согоян Н.С., Козаченко И.Ф., Адамян Л.В. Роль АМГ в репродуктивной системе женщин (обзор литературы). Проблемы репродукции. 2017;23(1):37 42.
- Castresana J. Genes on human chromosome 19 show extreme divergence from the mouse orthologs and a high GC content. Nucleic Acids Res. 2002 Apr 15;30(8):1751-6. doi: 10.1093/nar/30.8.1751. PMID: 11937628; PMCID: PMC113201.
- Dewailly D, Robin G, Peigne M, Decanter C, Pigny P, Catteau-Jonard S. Interactions between androgens, FSH, anti-Müllerian hormone and estradiol during folliculogenesis in the human normal and polycystic ovary. Hum Reprod Update. 2016 Nov;22(6):709-724. doi: 10.1093/humupd/dmw027. Epub 2016 Aug 27. PMID: 27566840.
- Didier Dewailly, Geoffroy Robin, Maëliss Peigne, Christine Decanter, Pascal Pigny, Sophie Catteau-Jonard, Interactions between androgens, FSH, anti-Müllerian hormone and estradiol during folliculogenesis in the human normal and polycystic ovary, Human Reproduction Update, Volume 22, Issue 6, 20 November 2016, Pages 709–724, https://doi.org/10.1093/humupd/dmw027
- Moolhuijsen LME, Visser JA. Anti-Müllerian hormone and ovarian reserve: update on assessing ovarian function. J Clin Endocrinol Metab 2020; 105 (11) 3361-3373
- Nilsson E, Rogers N, Skinner MK. Actions of anti-Mullerian hormone on the ovarian transcriptome to inhibit primordial to primary follicle transition. Reproduction. 2007 Aug;134(2):209-21. doi: 10.1530/REP-07–0119. PMID: 17660231; PMCID: PMC8260025.
- Rosenfield R.L., Wroblewski K., Padmanabhan V., Littlejohn E., Mortensen M., Ehrmann D.A. Anti-Müllerian hormone levels are independently related to ovarian hyperandrogenism and polycystic ovaries. Fertil. Steril. 2012; 98(1): 242-9.
- Skałba P, Cygal A, Dabkowska-Huć A. Wpływ hormonu anty-Mullerowskiego (AMH) na folikulogeneze [The influence of anti-Mullerian hormone on folliculogenesis]. Ginekol Pol. 2008 Feb;79(2):137-40. Polish. PMID: 18510094.
Статья поступила в редакцию 21 июля 2025 г.
Принята к печати 10 сентября 2025 г.
Received July 21, 2025
Accepted 10, September, 2025