Введение
Фолликул яичника - это основная структурная и функциональная единица яичника. В фолликуле содержится ооцит 1-го порядка, окруженный слоем гликопротеидов, которые в свою очередь окружены слоем гранулезных клеток. Клетки гранулезы являются важным соматическим компонентом яичника. Два разных типа гранулезных клеток, муральные гранулезные клетки и гранулезные клетки кумулюса, выполняют разные функции во время фолликулогенеза. Муральные гранулезные клетки вырабатывают эстроген во время фолликулярной фазы и прогестерон после овуляции, в то время как гранулезные клетки кумулюса плотно окружают ооцит и образуют кумулюс оофорус и внутренний слой клеток лучистого венца. Гранулезные клетки участвуют в двунаправленном обмене метаболитами с ооцитом, поскольку они образуют щелевые контакты, которые имеют решающее значение как для правильного созревания ооцита, так и для пролиферации гранулезных клеток. Взаимодействие между гранулезными клетками и ооцитами имеет решающее значение для скоординированного созревания ооцитов (Adriaenssens T, et al., 2011; Dompe, C., et al., 2021). Межклеточное взаимодействие между ооцитом и гранулезными клетками может быть опосредовано паракринной, аутокринной или эндокринной сигнализацией и отвечает за метаболическое взаимодействие, которое включает перенос глюкозы, нуклеотидов, аминокислот и метаболитов к яйцеклетке. Тесная взаимосвязь между ооцитом, гранулезными клетками и клетками-предшественниками поддерживает развитие фолликулов и созревание ооцитов посредством биосинтеза стеролов, регуляции мейоза, транскрипции генов и защиты ооцитов (Albertini D, et al., 2000; Assou S, et al.,2006; Tong, X. et al., 2014).
МикроРНК (miRNA) универсальные регуляторы экспрессии генов, они комплементарно спариваются с участками мРНК и ингибируют их трансляцию (Bartel, D.2004). Они связываются с 3'-нетранслируемой областью (3'-UTR) мРНК и вызывают ингибирование трансляции или деградацию мРНК, комплементарное связывание с соответствующими мРНК, ведет к ингибированию трансляции и деградации мРНК, и приводит к прекращению дальнейшей трансляции белка (Ambros, V. 2004). Известно, что таким способом микроРНК регулируют более 60% генов человека В геноме человека закодировано несколько тысяч микро РНК, образующих регуляторные сети, участвующие в сигнальных путях и различных клеточных процессах miRNA играет важнейшую регуляторную роль на разных стадиях онтогенеза, в норме и патологии (Griffiths-Jones, S., 2006; Аушев В., 2015; Кучер А., Бабушкина Н., 2011; Kozomara, A., 2019; Мустафин, Р.,Хуснутдинова, Э. 2025). В редких случаях микроРНК также могут положительно влиять на трансляцию и экспрессию генов (Sekar et al., 2016, Saravanan et al., 2015). С функциональной точки зрения отдельная миРНК так же важна, как и фактор транскрипции, поскольку она способна регулировать экспрессию множества генов-мишеней (Kozomara, A., 2019; Shang R. et al., 2023). Таким образом, микроРНК могут быть ключевыми регуляторами развития, физиологии и заболеваний (Erson, A., Petty, E.,2008; Sayed, D.,Abdellatif, M.,2011; Searles, C.2024; Cui, Y., et al.,2024; Омельчук, Е., и др., 2024). Несмотря на то, что основные особенности биогенеза и функционирования миРНК были открыты боле 20 лет назад, в последние годы продолжает раскрываться фундаментальная информация о структурной и молекулярной динамике основного механизма miRNA, о том, как субстраты и мишени miRNA отбираются из транскриптома, о новых путях многоуровневой регуляции биогенеза miRNA и механизмах взаимодействий miRNA, о их новой роли в регуляции клеточных процессов в онтогенезе.
Целью настоящего исследования было изучение транскриптомного профиля miRNA в кумулюсных гранулезных клетках человека.
Материал и методы
В полногеномном транскриптомном профилировании были использованы биологические образцы 5 женщин в возрасте от 26 до 34 лет: Все женщины были здоровы и включены в программу экстракорпорального оплодотворения по фактору мужского бесплодия. Для всех пациенток во время овариальной стимуляции использовали короткий протокол с антагонистами гонадотропинрелизинг гормона (Цетротид, Zentaris GmbH, Германия). Контролируемую гиперстимуляцию яичников вызывали введением рекомбинантного фолликулостимулирующего гормона человека (Гонал- ф Merck Serono, Италия) и человеческого менопаузального гонадотропина (Мериоферт, IBSA, Швейцария) в индивидуальных дозах гонадотропина. Овуляцию индуцировали хорионическим гонадотропином (Московский эндокринный завод ФГУП, Россия) для запуска созревания ооцитов на 9-12 день после начала гормональной стимуляции яичников. Объектами исследования были образцы гранулезных клеток кумулюса, полученные во время пункции фолликулов во время процедуры ЭКО в Центре репродукции человека и ЭКО (Ростов-на-Дону). Библиотеки для секвенирования микроРНК были подготовлены с использованием набора для подготовки библиотеки РНК NEBNext Small для Illumina. Секвенирование РНК проводилось на платформе Miseq (Illumina). Контроль качества секвенирования проводился с использованием программ FastQC (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/), удаление адаптеров проводили при помощи программы cutadapt (https://cutadapt.readthedocs.io/en/stable/). Было выполнено картирование и подсчет количества прочтений с использованием программы STAR 2.7.9a с параметром outFilterMismatchNmax. Далее для сборки транскриптов и оценки их уровня экспрессии использовался Cufflinks (https://cole-trapnell-lab.github.io/cufflinks). Экспрессию генов определяли количественно с использованием RPKM (reads per kilobase per million mapped reads — количество прочтений на килобазу на картированные риды) и показатель tpm (transcripts per million). На основании количества прочтений был установлен относительный уровень экспрессии микроРНК.
В работе использованы базы данных, которые находится в открытом доступе miRBase (http://mirbase.org), miRDB (www.mirdb.org) и miRTarBase (mirtarbase.mbc.nctu.edu.tw).
Результаты исследований
В общей сложности в транскриптом анализе мы исследовали 2653 микро РНК, в гранулезных клетках человека. Была обнаружена транскрипция 247 микро РНК. Учитывая, что количество сопоставленных прочтений зависит не только от уровня его экспрессии и длины гена, но и от глубины секвенирования, для нормализации этих зависимостей мы использовали показатель RPKM и показатель tpm (transcripts per million). TPM отражают относительную распространённость транскрипта среди популяции секвенированных транскриптонов и зависит от состава популяции РНК в образце.
Из 247 транскриптов микро РНК в гранулезных клетках кумулюса мы представляем 30 микро РНК с максимальными значениями tpm, как наиболее значимые (таблица 1). Как видно из результатов исследований транскриптомный профиль зрелых miRNA гранулезных клеток человека в кумулюсе (без учета родственных последовательностей) представлен следующими молекулами: hsa-miR-320; hsa-miR-21; hsa-miR-193; hsa-miR-1307; hsa-miR-10395; hsa-miR-127; hsa-let-7; hsa-miR-99; hsa-miR-744 hsa-miR-514; hsa-miR-629; hsa-miR 125; hsa-miR 140; hsa-miR 149; hsa-miR 181; hsa-miR-1291; hsa-miR 125; hsa-miR 151; hsa-miR 106; hsa-miR 365. В этом перечне мы не уточняем зрелые мРНК которые различаются только в одной или двух позициях, и им присваиваются буквенные суффиксы (например hsa-miR-320b и hsa-miR-320c), и когда две разные зрелые последовательности микроРНК вырезаются из противоположных концов одного и того же предшественника шпильки и обозначаются hsa-miR-193b-3p(3′-конец) и hsa-miR-193b-5p (5′-конец). В таблице 1 представлены уникальные транскрибирующиеся miRNA по номенклатуре ID miRBase (http://mirbase.org).
Таблица 1
Транскриптомный профиль miRNA в кумулюсных гранулезных клетках человека.
|
№ |
ID (miRBase) |
tpm |
|
1. |
hsa-miR-320b |
15679,8 |
|
2. |
hsa-miR-320c |
5945,8 |
|
3. |
hsa-miR-21-5p |
2893,4 |
|
4. |
hsa-miR-193b-3p |
1842,2 |
|
5. |
hsa-miR-1307-3p |
1721,9 |
|
6. |
hsa-miR-10395-5p |
1623,7 |
|
7. |
hsa-miR-320a-3p |
1576,7 |
|
8. |
hsa-miR-127-3p |
1514,5 |
|
9. |
hsa-miR-210-3p |
1508,9 |
|
10. |
hsa-let-7a-5p |
1208,8 |
|
11. |
hsa-let-7b-5p |
1085,7 |
|
12. |
hsa-miR-99a-5p |
1085,7 |
|
13. |
hsa-miR-744-5p |
915,5 |
|
14. |
hsa-miR-514a-3p |
874,1 |
|
15. |
hsa-miR-629-5p |
803,6 |
|
16. |
hsa-miR-125b-5p |
773,1 |
|
17. |
hsa-miR-140-3p |
759,3 |
|
18. |
hsa-miR-149-5p |
721,9 |
|
19. |
hsa-miR-181a-5p |
674,9 |
|
20. |
hsa-miR-1291 |
603,0 |
|
21. |
hsa-miR-125a-5p |
569,8 |
|
22. |
hsa-miR-99b-5p |
560,1 |
|
23. |
hsa-miR-151a-3p |
539,4 |
|
24. |
hsa-miR-106b-3p |
524,2 |
|
25. |
hsa-miR-365a-5p |
449,5 |
|
26. |
hsa-miR-10395-3p |
432,9 |
|
27. |
hsa-miR-99b-3p |
427,4 |
|
28. |
hsa-miR-193b-5p |
421,8 |
|
29. |
hsa-let-7c-5p |
396,9 |
|
30. |
hsa-miR-3615 |
387,3 |
Как видно из представленных данных наибольшее число транскрибирующихся копий в гранулезных клетках кумулюса приходится на hsa-miR-320b (15679,8) и hsa-miR-320c (5945,8) и hsa-miR-320a-3p (1576,7). Без учета родственных последовательностей hsa-miR-320 более 23200 tmp. Родственные последовательности hsa-miR-320 локализованы на разных хромосомах и отличаются друг от друга одним нуклеотидом (таблица 2).
Таблица 2
Локализация и последовательности зрелых микро РНКhsa-miR-320
|
ID (miRBase) |
Последовательность |
Хромосома |
|
hsa-miR-320с |
AAAAGCUGGGUUGAGAGGGU |
18
|
|
hsa-miR-320b
|
AAAAGCUGGGUUGAGAGGGCAA |
1 |
|
hsa-miR-320a-3p |
AAAAGCUGGGUUGAGAGGGCGA |
8
|
Поскольку одна микроРНК может воздействовать на несколько генов, а один ген может быть мишенью для нескольких микроРНК, мы использую базу данных MicroRNA Target Prediction Database (miRDB, www.mirdb.org), которая была разработана на основе анализа тысяч взаимодействий микроРНК с мишенями в ходе высокопроизводительных экспериментов по секвенированию, стремились выявить любые общие пути взаимодействия hsa-miR-320 с белками мишенями. В miRDB предсказано 1045 мишеней для hsa-miR-320b. В том числе hsa-miR-320b является мишенью для 35 белков с различными типами цинковых пальцев, 27 факторов и субьединиц транскрипционных комплексов, 27 различных киназ, 17 мембранных белков, 5 циклинзависимых киназ, 5 факторов апоптоза и др. (https://mirdb.org/cgi-bin/search.cgi?searchType=miRNA&searchBox=hsa-miR-320b&full=1). Для понимания в каких клеточных процессах принимает участие hsa-mir-320b , на основе данных полученных в 147 статьях, которые представлены в открытом доступе и упоминают hsa-mir-320b создано «облако слов» (рис.1).
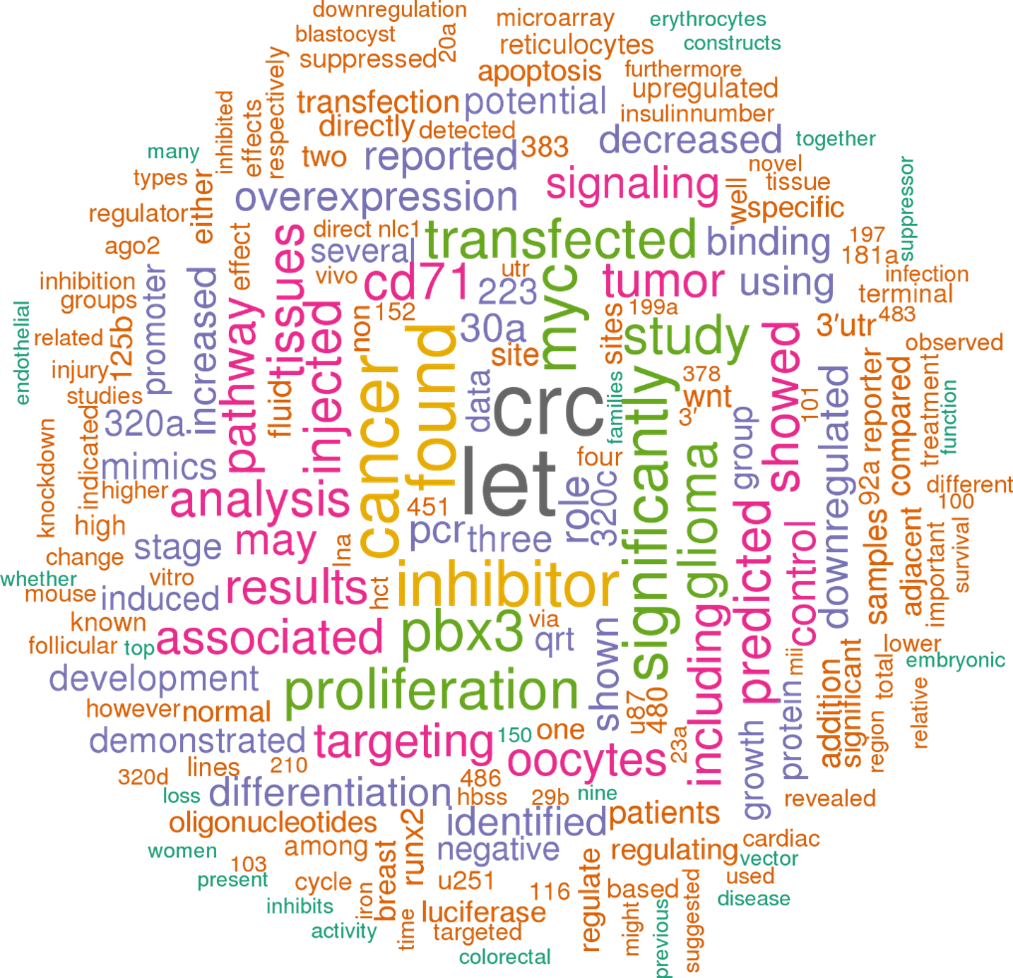
Рис.1. «Облако слов» созданное на основе 147 статей, которые представлены в открытом доступе и упоминают hsa-mir-320b (https://www.mirbase.org/hairpin/MI0003776).
Как видно из представленного «облака слов» hsa-mir-320b принимает участие в пролиферации, в сигнальных путях, в том числе и пути WNT, наиболее часто встречается в работах где исследуется процессы онкогенеза, в частости колоректальный рак (CRC), есть единичные работы о его роли в функционировании яичника и фолликула, анализ работ показывает, что достаточно часто hsa-mir-320b экспрессируется с родственными последоватльностями 320а, 320d.
Ранее было показано, что результате наиболее распространёнными микроРНК в кумулюсных клетках оказались let-7b, let-7c и hsa-mir-21 (Assou, S.;2014). В наших исследованиях транскптома гранулезных клеток кумулюса также отмечена их значительная экспрессия (таблица 1). hsa-miR-21-3p локализована на 17 хромосоме и для нее предсказано 595 мишеней (https://www.mirbase.org/results/?query=hsa-miR-21). «Облако слов» созданное на основе 1559 статей, в которых исследована hsa-mir-21, показывает, что эта молекула играет решающую роль в регуляции апоптоза и роста организмов, а также ассоциирована с подавлением опухолей (рис.2). В единичных работах приводятся данные о том , что miR-21-3p влияет на аутофагию гранулезных клеток (у крупного рогатого скота), как показали Ма и др., воздействуя на VEGFA и ослабляя передачу сигналов PI3K/AKT, что приводит к ингибированию аутофагии (Ma, L., 2019).
Семейство let-7 - это высококонсервативное семейство микроРНК, играющее важную роль в развитии, дифференцировке, метаболизме, пролиферации и апоптозе у многих видов. МикроРНК let-7, впервые обнаруженная у нематоды C. elegans, является ключевым регулятором развития и участвует в регуляции экспрессии генов на посттранскрипционном уровне. Семейство let-7 у человека состоит из 13 членов (Roush and Slack, 2008). Семейство hsa-let-7 дифференциально экспрессируется во время атрезии фолликулов (Su, J et al., 2012: Gong Z. et al., 2019). уровни экспрессии генов mir-let-7a, let-7b, let-7c и let-7i были снижены в ранних и прогрессирующих атретических фолликулах по сравнению со здоровыми фолликулами (Zhou, j., 2015). показано, что let-7g регулирует апоптоз гранулезных клеток воздействуя на TGFBR1 и подавляя сигнальный путь TGF-Β ( J Zhou, 2015).

Рис.2. «Облако слов» созданное на основе 1559 статей, в которых исследована hsa-mir-21 (https://www.mirbase.org/hairpin/MI0000077).
Сверхэкспрессия mir-let-7g повышала уровень апоптоза в гранулезных клетках мышей и экспрессию FOXO1 в гранулезных клетках, что привело к накоплению дефосфорилированного FOXO1 в ядре (Cao R.,et al., 2015). В таблице 3 Транскриптомный профиль 11 микроРНК семейства hsa-let-7, который был определен в наших исследованиях в кумулюсных гранулезных клетках человека. Повышенная экспрессия miRNA let-7 в фолликулярном микроокружении яичников и влияет на экспрессию рецептора инсулиноподобного фактора роста 1-го типа (IGF1R) в клетках гранулезы, нарушаяя их пролиферацию, уровень секреции стероидных гормонов, энергетический метаболизм аденозинтрифосфата и уровень окислительного стресса, вызванного активными формами кислорода (Shi, L.,2025). Также показано, что hsa-let-7 регулирует экспрессию рецептора эстрогена (Zhang, R.,2012). Транскриптомный профиль семейства hsa-let-7a в кумулюсных гранулезных клетках человека представлен в таблице 3.
Таблица 3
Транскриптомный профиль семейства hsa-let-7a в кумулюсных гранулезных клетках человека.
|
№ |
ID (miRBase) |
tpm |
|
1 |
hsa-let-7a-5p |
1208,8 |
|
3 |
hsa-let-7c-5p |
396,9 |
|
4 |
hsa-let-7g-5p |
160,4 |
|
5 |
hsa-let-7f-5p |
157,7 |
|
6 |
hsa-let-7i-5p |
117,6 |
|
7 |
hsa-let-7e-5p |
33,2 |
|
8 |
hsa-miR-7-5p |
22,1 |
|
9 |
hsa-let-7d-5p |
13,8 |
|
10 |
hsa-let-7d-3p |
2,7 |
|
11 |
hsa-let-7b-3p |
1,4 |
Как видно из таблицы 3 из 13 членов семейства hsa-let-7, 11 транскриптов микро РНК было обнаружено в гранулезных клетках кумулюса здоровых женщин. Для hsa-let-7a-5p в каталоге miRDB предсказано 990 мишеней. Из них 15 белков различных рецепторов, 9 белков -факторов роста а, 8 факторов транскрипции, 8 различных семейств белков переносчиков,5 белков в различных гомеобксах. На рисунке 3 представлено «облако слов» биологических процессов, созданное на основе 1270 статей, в которых исследована hsa-let-7а (https://www.mirbase.org)
В работах китайских ученых была исследована роль микроРНК в клетках гранулезы яичников в норме и при заболеваниях. Ими отмечено 10 наиболее распространённых микроРНК в муральных гранулезных клетках и гранулезных клетках кумулюса: miR-21-5p, let-7a-5p, let-7f-5p, miR-26a-5p, let-7b-5p, let-7g-5p, miR-103a-3p, miR-125a-5p, miR-92a-3p, miR-320a и другие микроРНК (Tu, J.,2019). Как видно из таблицы1 микро РНК семейства let-7 miR-21-5p, miR-320a miR-125a-5p имеют пересечение с траскриптомным профилем микро РНК в гранулезных клетках полученном в нашем исследовании и исследовании китайских ученых.

Рис. 3. «Облако слов» биологических процессов, созданное на основе1270 статей, в которых исследована hsa-let-7а (https://www.mirbase.org)
Заключение.
Многие микроРНК экспрессируются в гранулезных клетках и напрямую регулируют нормальное развитие и функционирование фолликулов яичников. Одна микроРНК может подавлять сотни, даже тысячи генов, а один ген может модулироваться несколькими микроРНК. Наше исследование позволяет получить ценные сведения о транскриптоме и регуляторных механизмах транскрипции в ооцитах и клетках кумулюса. В этом исследовании мы оценивали только транскрипционный профиль клеток кумулюса, непосредственно окружающих ооцит. Эти клетки кумулюса играют ключевую роль на заключительных стадиях созревания ооцита, напрямую контактируя с ооцитом и доставляя метаболические субстраты. Выявление микроРНК, специфичных для гранулезных клеток, поможет исследователям лучше понять механизмы, лежащие в основе заболеваний яичников.
Финансирование
Исследование выполнено при финансовой поддержке Российского научного фонда, проект № 23–15-00464.
Список литературы
- Аушев В. Н. МикроРНК: малые молекулы с большим значением //Клиническая онкогематология. Фундаментальные исследования и клиническая практика. – 2015. – Т. 8. – №. 1. – С. 1-12.
- Кучер А. Н., Бабушкина Н. П. Роль микро-РНК, генов их биогенеза и функционирования в развитии патологических состояний у человека //Медицинская генетика. – 2011. – Т. 10. – №. 1. – С. 3-13.
- Мустафин, Р. Н., & Хуснутдинова, Э. К. (2025). Взаимосвязь мобильных генетических элементов с некодирующими РНК в развитии атеросклероза (обзор). Научные результаты биомедицинских исследований, 11(1), 31-56.
- Омельчук, Е. П., Тимошкина, Н. Н., Росторгуев, Э. Е., & Дженкова, Е. А. (2024). Циркулирующие биомаркеры глиом (обзор литературы). Клиническая лабораторная диагностика, 69(8), 411-20.
- Albertini DF, Combelles CM, Benecchi E, Carabatsos MJ (2000) Cellular basis for paracrine regulation of ovarian follicle development. Reproduction 121: 647–653.
- Ambros, V. (2004). The functions of animal microRNAs. Nature, 431(7006), 350-355.
- Assou S, Anahory T, Pantesco V, Le Carrour T, Pellestor F, et al. (2006) The human cumulus – oocyte complex gene-expression profile. Hum Reprod 21: 1705–1719.
- Assou, S.; Al-Edani, T.; Haouzi, D.; Philippe, N.; Lecellier, C.H.; Piquemal, D.; Commes, T.; Aït-Ahmed, O.; Dechaud, H.; Hamamah, S. MicroRNAs: New candidates for the regulation of the human cumulus-oocyte complex. Reprod. 2013, 28, 3038–3049.
- Adriaenssens T, Segers I, Wathlet S, Smitz J (2011) The cumulus cell gene expression profile of oocytes with different nuclear maturity and potential for blastocyst formation. J Assist Reprod Genet 28: 31–40.
- Bartel, D. P. (2004). MicroRNAs: genomics, biogenesis, mechanism, and function. cell, 116(2), 281-297.
- Cao R., Wu, W. J., Zhou, X. L., Xiao, P., Wang, Y., & Liu, H. L. Expression and preliminary functional profiling of the let-7 family during porcine ovary follicle atresia //Molecules and cells. – 2015. – Т. 38. – №. 4. – С. 304-311.
- Cheng, Y., Liu, X., Zhang, S., Lin, Y., Yang, J., & Zhang, C. (2009). MicroRNA-21 protects against the H2O2-induced injury on cardiac myocytes via its target gene PDCD4. Journal of molecular and cellular cardiology, 47(1), 5-14.
- Cui, Y., Qi, Y., Ding, L., Ding, S., Han, Z., Wang, Y., & Du, P. (2024). miRNA dosage control in development and human disease. Trends in Cell Biology, 34(1), 31-47.
- Dompe, C., Kulus, M., Stefańska, K., Kranc, W., Chermuła, B., Bryl, R., ... & Kempisty, B. (2021). Human granulosa cells—stemness properties, molecular cross-talk and follicular angiogenesis. Cells, 10(6), 1396.
- Erson, A. E., & Petty, E. M. (2008). MicroRNAs in development and disease. Clinical genetics, 74(4), 296-306
- Gong, Z., Yang, J., Bai, S., & Wei, S. (2019). MicroRNAs regulate granulosa cells apoptosis and follicular development—A review. Asian-Australasian journal of animal sciences, 33(11), 1714.
- Griffiths-Jones, S., Grocock, R. J., Van Dongen, S., Bateman, A., & Enright, A. J. (2006). miRBase: microRNA sequences, targets and gene nomenclature. Nucleic acids research, 34(suppl_1), D140-D144.
- Kozomara, A., Birgaoanu, M., & Griffiths-Jones, S. (2019). miRBase: from microRNA sequences to function. Nucleic acids research, 47(D1), D155-D162.
- Ma, L., Zheng, Y., Tang, X., Gao, H., Liu, N., Gao, Y., ... & Jiang, Z. (2019). miR-21-3p inhibits autophagy of bovine granulosa cells by targeting VEGFA via PI3K/AKT signaling. Reproduction, 158(5), 441-452.
- Roush S., Slack F. J. The let-7 family of microRNAs //Trends in cell biology. – 2008. – Т. 18. – №. – С. 505-516.
- Searles, C. D. (2024). MicroRNAs and cardiovascular disease risk. Current Cardiology Reports, 26(2), 51-60.
- Sayed, D., & Abdellatif, M. (2011). MicroRNAs in development and disease. Physiological reviews, 91(3), 827-887.
- Sekar, D., Venugopal, B., Sekar, P., & Ramalingam, K. (2016). Role of microRNA 21 in diabetes and associated/related diseases. Gene, 582(1), 14-18.
- Shang, R., Lee, S., Senavirathne, G., & Lai, E. C. (2023). microRNAs in action: biogenesis, function and regulation. Nature Reviews Genetics, 24(12), 816-833.
- Shi, L., Ying, H., Dai, Y., Rong, Y., Chen, J., Zhou, F., ... & Zhang, S. (2025). Upregulated let-7 expression in the follicular fluid of patients with endometriomas leads to dysfunction of granulosa cells through targeting of IGF1R. Human Reproduction, 40(1), 119-137.
- Su, J. L., Chen, P. S., Johansson, G., & Kuo, M. L. (2012). Function and regulation of let-7 family microRNAs. Microrna, 1(1), 34-39.
- Tong, X. H., Xu, B., Zhang, Y. W., Liu, Y. S., & Ma, C. H. (2014). Research resources: comparative microRNA profiles in human corona radiata cells and cumulus oophorus cells detected by next-generation small RNA sequencing. PLoS One, 9(9), e106706.
- Tu, J., Cheung, A. H. H., Chan, C. L. K., & Chan, W. Y. (2019). The role of microRNAs in ovarian granulosa cells in health and disease. Frontiers in endocrinology, 10, 174.
- Zhang, R., He, Y., Zhang, X., Xing, B., Sheng, Y., Lu, H., & Wei, Z. (2012). Estrogen receptor-regulated microRNAs contribute to the BCL2/BAX imbalance in endometrial adenocarcinoma and precancerous lesions. Cancer letters, 314(2), 155-165.
- Zhou, J., Liu, J., Pan, Z., Du, X., Li, X., Ma, B., ... & Liu, H. (2015). The let-7g microRNA promotes follicular granulosa cell apoptosis by targeting transforming growth factor-β type 1 receptor. Molecular and Cellular Endocrinology, 409, 103-112.
Статья поступила в редакцию 18 марта 2025 г.
Принята к печати 28 марта 2025 г.
Received 18, March, 2025
Accepted 28, March, 2025